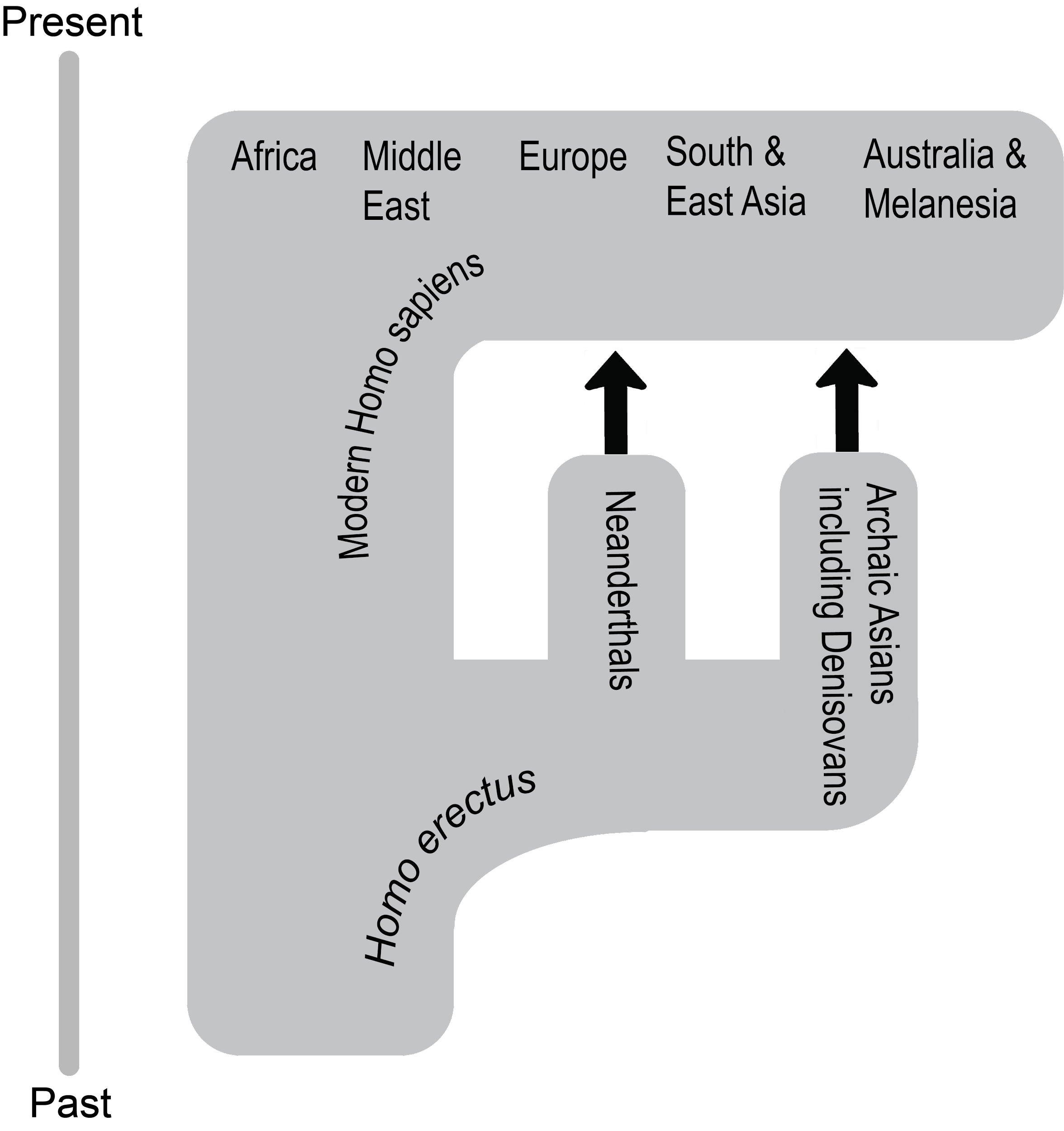
13 Modern Homo sapiens
Keith Chan, Ph.D., Grossmont-Cuyamaca Community College District and MiraCosta College
Student contributors to this chapter: Lily Berruyer, Lyn Loytchenko, and Sarah Cupidio
This chapter is a revision from “Chapter 12: Modern Homo sapiens” by Keith Chan. In Explorations: An Open Invitation to Biological Anthropology, first edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0.
Learning Objectives
- Identify the skeletal and behavioral traits that represent modern Homo sapiens.
- Critically evaluate different types of evidence for the origin of our species in Africa and our expansion around the world.
- Understand how the human lifestyle changed when people transitioned from foraging to agriculture.
- Hypothesize how human evolutionary trends may continue into the future.
The walls of a pink limestone cave in the hillside of Jebel Irhoud jutted out of the otherwise barren landscape of the Moroccan desert (Figure 13.1). Miners had excavated the cave in the 1960s, revealing some fossils. In 2007, a re-excavation of the site became a momentous occasion for science. A fossil cranium unearthed by a team of researchers was barely visible to the untrained eye. Just the fossil’s robust brows were peering out of the rock. This research team from the Max Planck Institute for Evolutionary Anthropology was the latest to explore the ancient human presence in this part of North Africa after a find by miners in 1960. Excavating near the first discovery, the researchers wanted to learn more about how Homo sapiens lived far from East Africa, where we thought our species originated.

The scientists were surprised when they analyzed the cranium, named Irhoud 10, and other fossils. Statistical comparisons with other human crania concluded that the Irhoud face shapes were typical of recent modern humans while the braincases matched ancient modern humans. Based on the findings of other scientists, the team expected these modern Homo sapiens fossils to be around 200,000 years old. Instead, dating revealed that the cranium had been buried for around 315,000 years.
Together, the modern-looking facial dimensions and the older date reshaped the interpretation of our species: modern Homo sapiens. Some key evolutionary changes from the archaic Homo sapiens (described in Chapter 11) to our species today happened 100,000 years earlier than we had thought and across the vast African continent rather than concentrated in its eastern region.
This revelation in the study of modern Homo sapiens is just one of the latest in this continually advancing area of biological anthropology. Researchers today are still discovering amazing fossils and ingenious ways to collect data and test hypotheses about our past. Through the collective work of many scientists, we are building an overall theory of modern human origins.
Defining Modernity
What defines modern Homo sapiens when compared to archaic Homo sapiens? Modern humans, like you and me, have a set of derived traits that are not seen in archaic humans or any other hominin. As with other transitions in hominin evolution, such as increasing brain size and bipedal ability, modern traits do not appear fully formed or all at once. In other words, the first modern Homo sapiens was not just born one day from archaic parents. The traits common to modern Homo sapiens appeared in a mosaic manner: gradually and out of sync with one another. There are two areas to consider when tracking the complex evolution of modern human traits. One is the physical change in the skeleton. The other is behavior inferred from the size and shape of the cranium and material culture evidence.
Skeletal Traits
The skeleton of modern Homo sapiens is less robust than that of archaic Homo sapiens. In other words, the modern skeleton is gracile, meaning that the structures are thinner and smoother. Differences related to gracility in the cranium are seen in the braincase, the face, and the mandible. There are also broad differences in the rest of the skeleton.
Cranial Traits
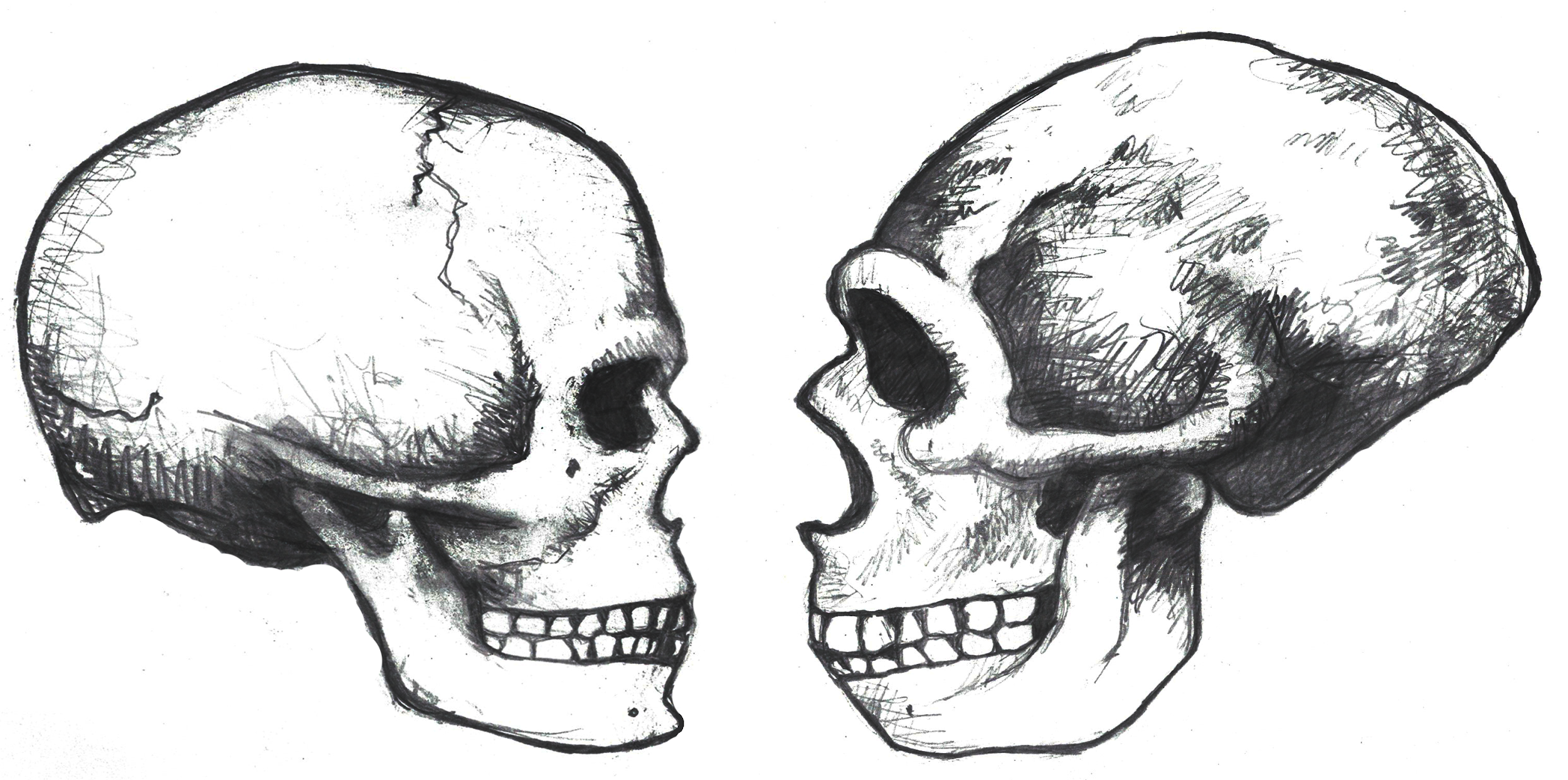
Several elements of the braincase differ between modern and archaic Homo sapiens. Overall, the shape is much rounder, or more globular, on a modern skull (Lieberman, McBratney, and Krovitz 2002; Neubauer, Hublin, and Gunz 2018; Pearson 2008; Figure 13.2). You can feel the globularity of your own modern human skull. Feel the height of your forehead with the palm of your hand. Viewed from the side, the tall vertical forehead of a modern Homo sapiens stands out when compared to the sloping archaic version. This is because the frontal lobe of the modern human brain is larger than the one in archaic humans, and the skull has to accommodate the expansion. The vertical forehead reduces a trait that is common to all other hominins: the brow ridge or supraorbital torus. The parietal lobes of the brain and the matching parietal bones on either side of the skull both bulge outward more in modern humans. At the back of the skull, the archaic occipital bun is no longer present. Instead, the occipital region of the modern human cranium has a derived tall and smooth curve, again reflecting the globular brain inside.
The trend of shrinking face size across hominins reaches its extreme with our species as well. The facial bones of a modern Homo sapiens are extremely gracile compared to all other hominins (Lieberman, McBratney, and Krovitz 2002). Continuing a trend in hominin evolution, technological innovations kept reducing the importance of teeth in reproductive success (Lucas 2007). As natural selection favored smaller and smaller teeth, the surrounding bone holding these teeth also shrank.
Related to smaller teeth, the mandible is also gracile in modern humans when compared to archaic humans and other hominins. Interestingly, our mandibles have pulled back so far from the prognathism of earlier hominins that we gained an extra structure at the most anterior point, called the mental eminence. You know this structure as the chin. At the skeletal level, it resembles an upside-down “T” at the centerline of the mandible (Pearson 2008). Looking back at archaic humans, you will see that they all lack a chin. Instead, their mandibles curve straight back without a forward point. What is the chin for and how did it develop? Flora Gröning and colleagues (2011) found evidence of the chin’s importance by simulating physical forces on computer models of different mandible shapes. Their results showed that the chin acts as structural support to withstand strain on the otherwise gracile mandible.
Postcranial Gracility
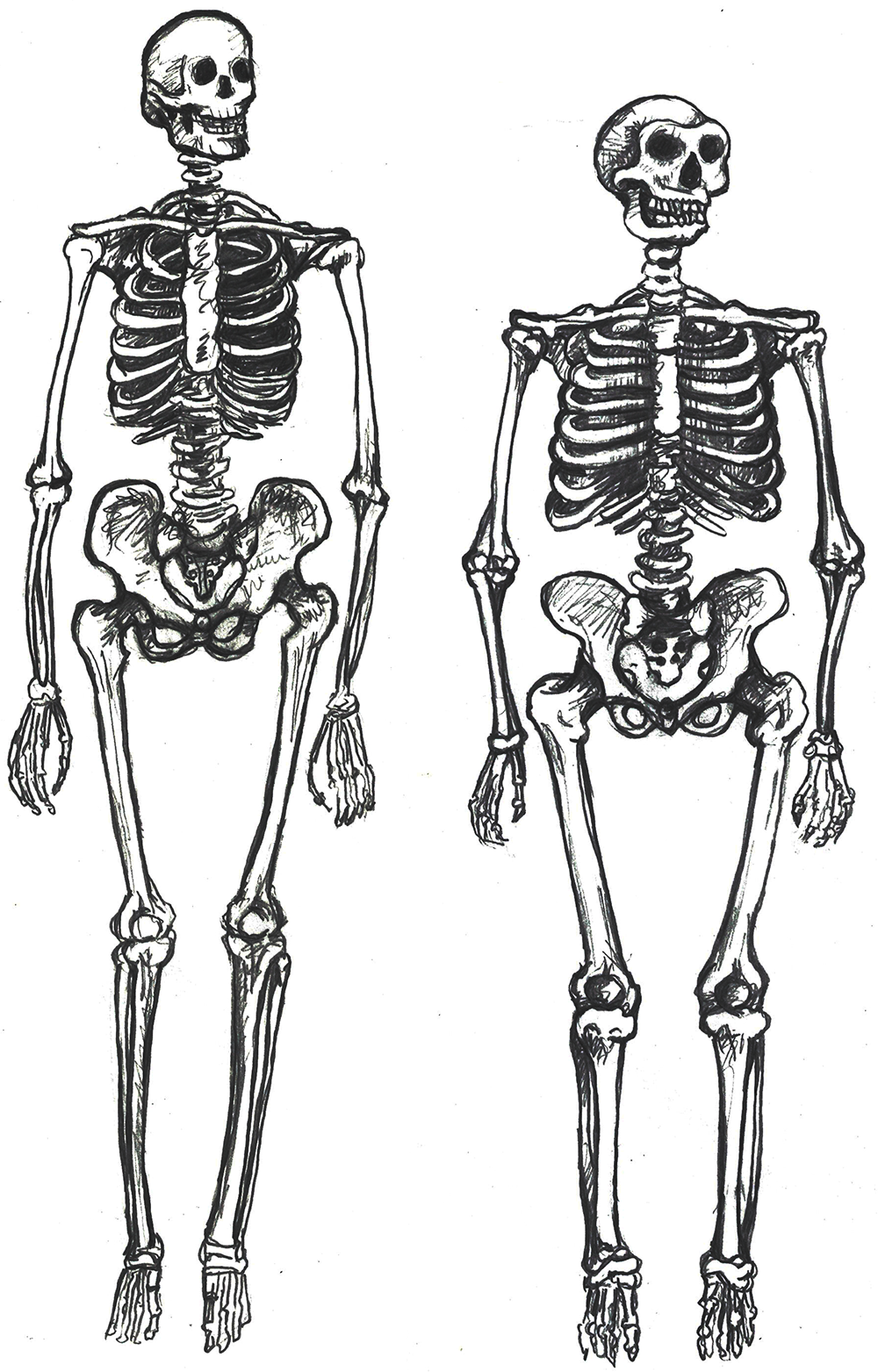
The rest of the modern human skeleton is also more gracile than its archaic counterpart. The differences are clear when comparing a modern Homo sapiens with a cold-adapted Neanderthal (Sawyer and Maley 2005), but the trends are still present when comparing modern and archaic humans within Africa (Pearson 2000). Overall, a modern Homo sapiens postcranial skeleton has thinner cortical bone, smoother features, and more slender shapes when compared to archaic Homo sapiens (Figure 13.3). Comparing whole skeletons, modern humans have longer limb proportions relative to the length and width of the torso, giving us lankier outlines.
Why is our skeleton so gracile compared to those of other hominins? Natural selection can drive the gracilization of skeletons in several ways (Lieberman 2015). A slender frame is believed to be adapted for the efficient long-distance running ability that started with Homo erectus. Furthermore, it is argued that slenderness is a genetic adaptation for cooling an active body in hotter climates, which aligns with the ample evidence that Africa was the home continent of our species.
Behavioral Modernity
Aside from physical differences in the skeleton, researchers have also uncovered evidence of behavioral changes associated with increased cultural complexity from archaic to modern humans. How did cultural complexity develop? Two investigations into this question are archaeology and the analysis of reconstructed brains.
Archaeology tells us much about the behavioral complexity of past humans by interpreting the significance of material culture. In terms of advanced culture, items created with an artistic flair, or as decoration, speak of abstract thought processes (Figure 13.4). The demonstration of difficult artistic techniques and technological complexity hints at social learning and cooperation as well. According to paleoanthropologist John Shea (2011), one way to track the complexity of past behavior through artifacts is by measuring the variety of tools found together. The more types of tools constructed with different techniques and for different purposes, the more modern the behavior. Researchers are still working on an archaeological way to measure cultural complexity that is useful across time and place.

The interpretation of brain anatomy is another promising approach to studying the evolution of human behavior. When looking at investigations on this topic in modern Homo sapiens brains, researchers found a weak association between brain size and test-measured intelligence (Pietschnig et al. 2015). Additionally, they found no association between intelligence and biological sex. These findings mean that there are more significant factors that affect tested intelligence than just brain size. Since the sheer size of the brain is not useful for weighing intelligence within a species, paleoanthropologists are instead investigating the differences in certain brain structures. The differences in organization between modern Homo sapiens brains and archaic Homo sapiens brains may reflect different cognitive priorities that account for modern human culture. As with the archaeological approach, new discoveries will refine what we know about the human brain and apply that knowledge to studying the distant past.
Taken together, the cognitive abilities in modern humans may have translated into an adept use of tools to enhance survival. Researchers Patrick Roberts and Brian A. Stewart (2018) call this concept the generalist-specialist niche: our species is an expert at living in a wide array of environments, with populations culturally specializing in their own particular surroundings. The next section tracks how far around the world these skeletal and behavioral traits have taken us.
First Africa, Then the World
What enabled modern Homo sapiens to expand its range further in 300,000 years than Homo erectus did in 1.5 million years? The key is the set of derived biological traits from the last section. It is theorized that the gracile frame and neurological anatomy allowed modern humans to survive and even flourish in the vastly different environments they encountered. Based on multiple types of evidence, the source of all of these modern humans was Africa. Instead of originating from just one location, evidence shows that modern Homo sapiens evolution occurred in a complex gene flow network across Africa, a concept called African multiregionalism (Scerri et al. 2018).
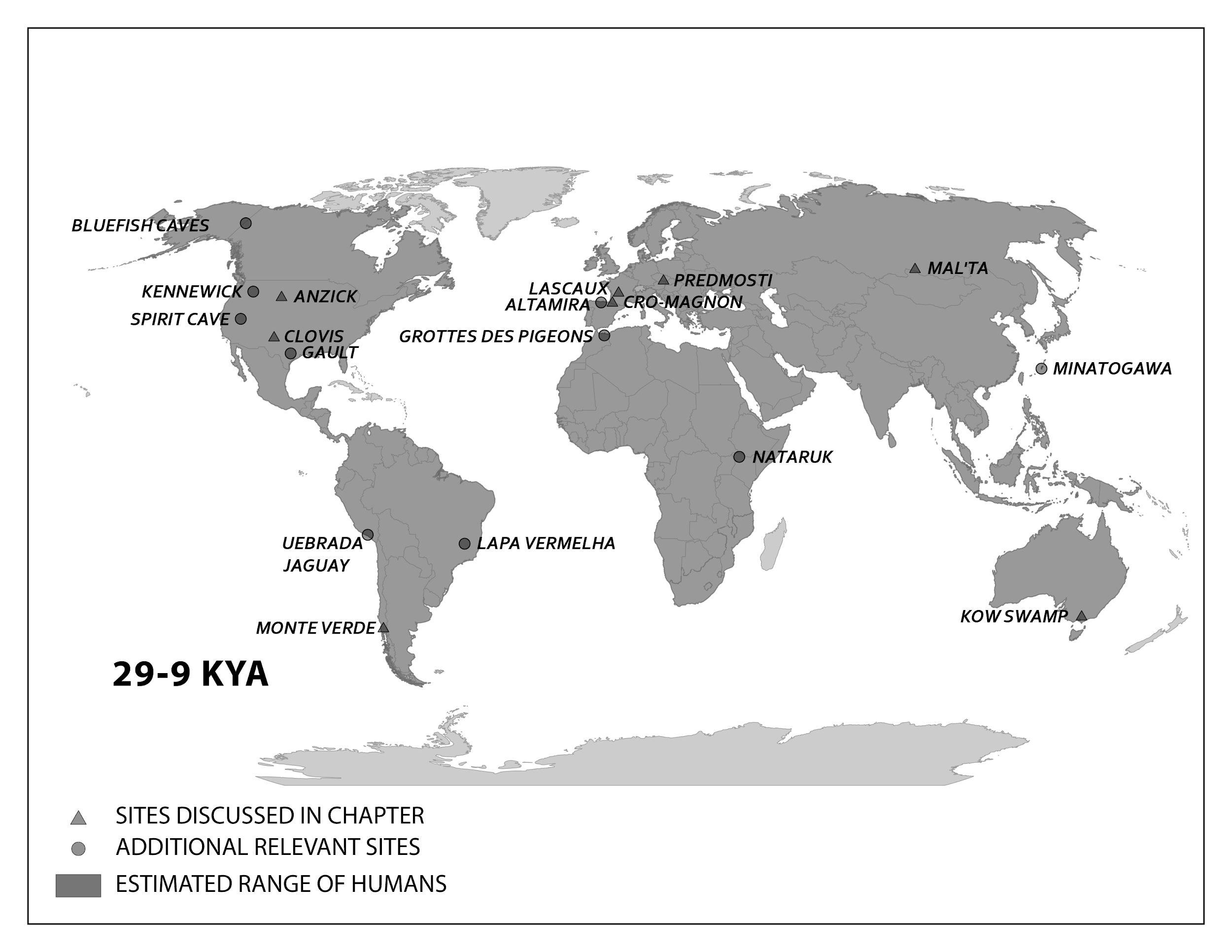
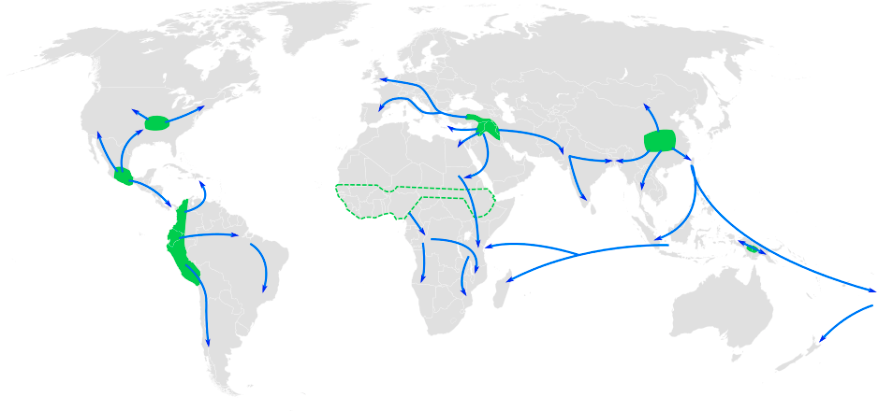
This section traces the origin of modern Homo sapiens and the massive expansion of our species across all of the continents (except Antarctica) by 12,000 years ago. While modern Homo sapiens first shared geography with archaic humans, modern humans eventually spread into lands where no human had gone before. Figure 13.5 shows the broad routes that our species took expanding around the world. I encourage you to make your own timeline with the dates in this part to see the overall trends.
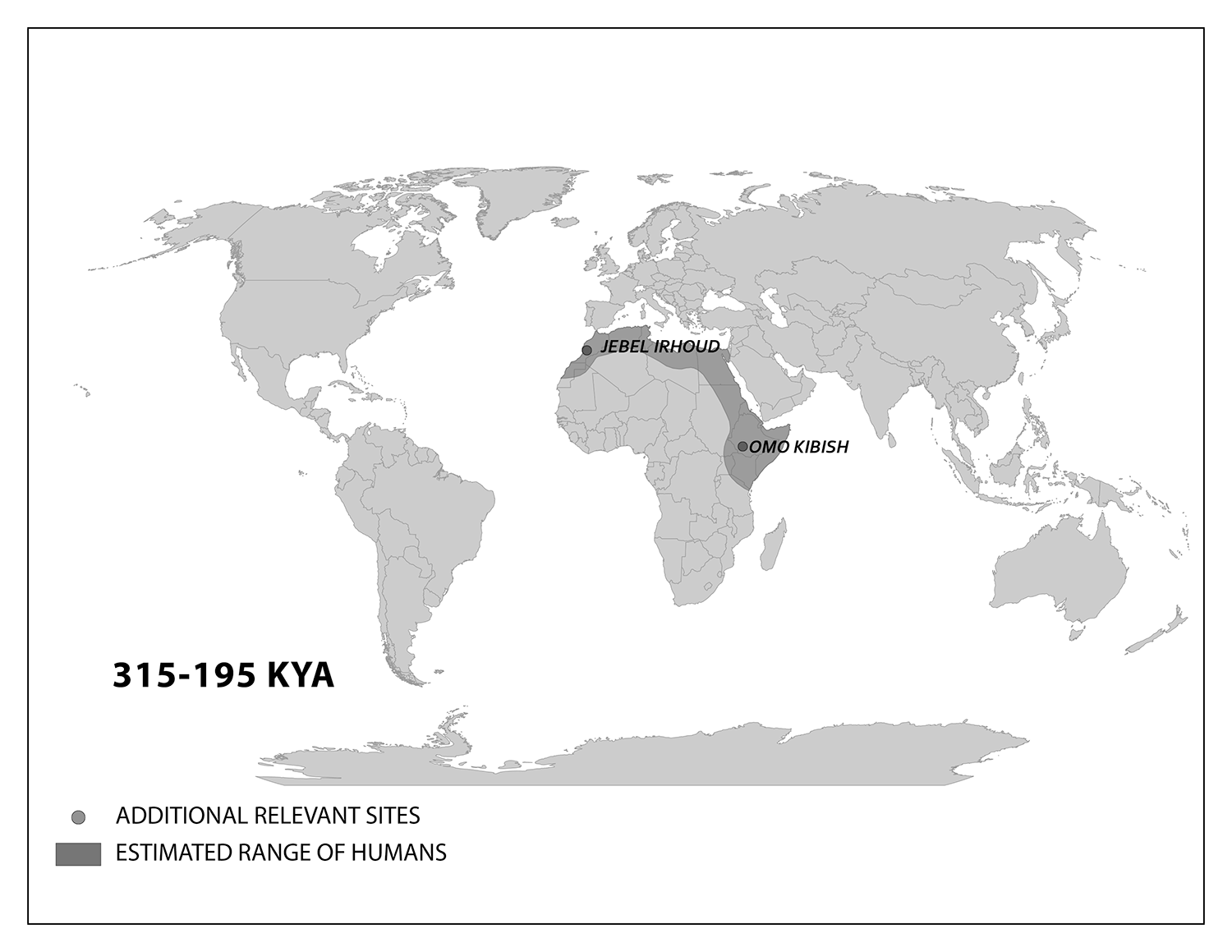
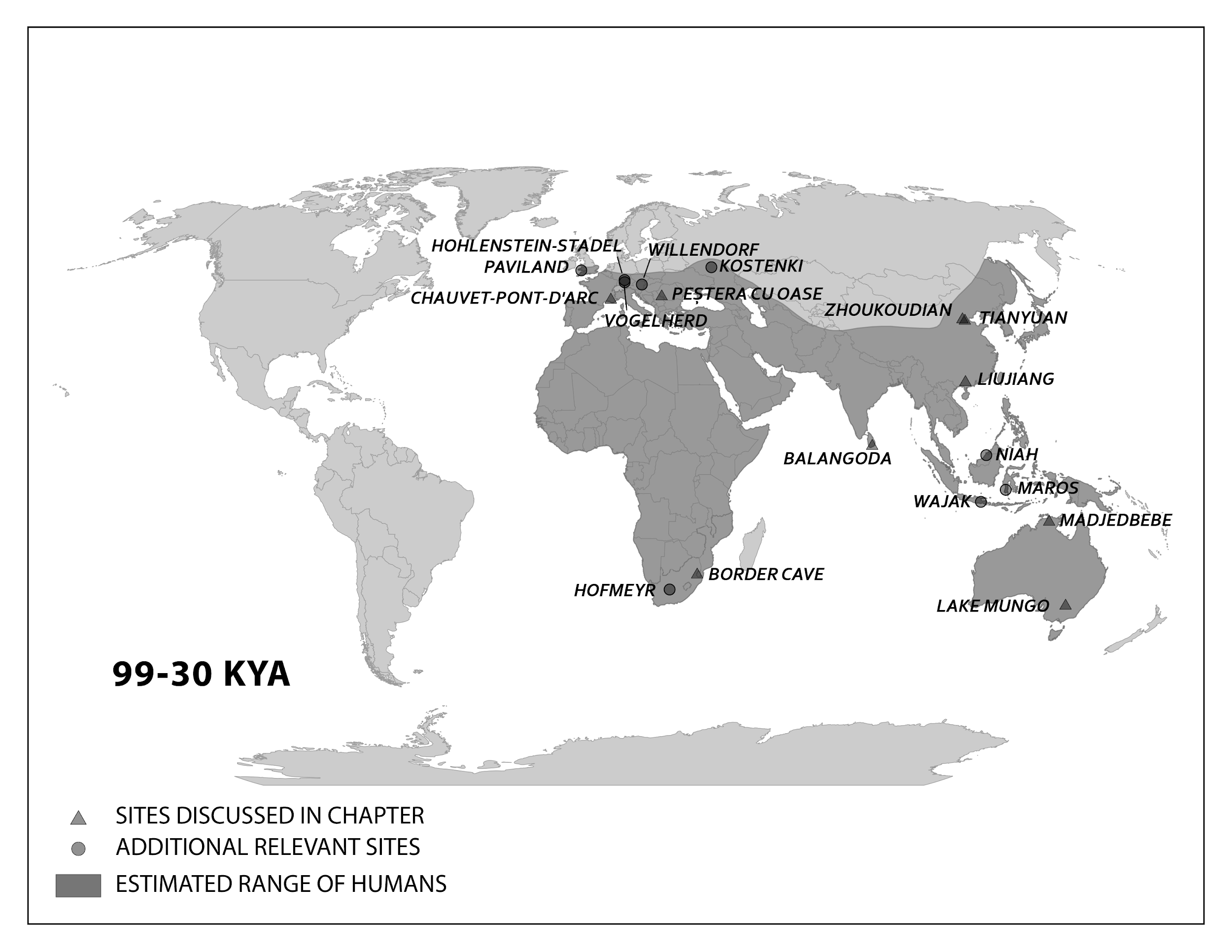
Modern Homo sapiens Biology and Culture in Africa
We start with the ample fossil evidence supporting the theory that modern humans originated in Africa during the Middle Pleistocene, having evolved from African archaic Homo sapiens. The earliest dated fossils considered to be modern actually have a mosaic of archaic and modern traits, showing the complex changes from one type to the other. Experts have various names for these transitional fossils, such as Early Modern Homo sapiens or Early Anatomically Modern Humans. However they are labeled, the presence of some modern traits means that they illustrate the origin of the modern type. Three particularly informative sites with fossils of the earliest modern Homo sapiens are Jebel Irhoud, Omo, and Herto.
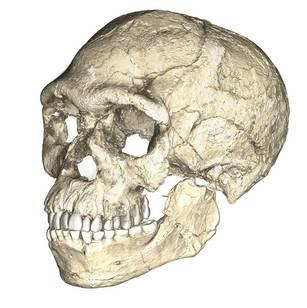
Recall from the start of the chapter that the most recent finds at Jebel Irhoud are now the oldest dated fossils that exhibit some facial traits of modern Homo sapiens. Besides Irhoud 10, the cranium that was dated to 315,000 years ago (Hublin et al. 2017; Richter et al. 2017), there were other fossils found in the same deposit that we now know are from the same time period. In total there are at least five individuals, representing life stages from childhood to adulthood. These fossils form an image of high variation in skeletal traits. For example, the skull named Irhoud 1 has a primitive brow ridge, while Irhoud 2 and Irhoud 10 do not (Figure 13.6). The braincases are lower than what is seen in the modern humans of today but higher than in archaic Homo sapiens. The teeth also have a mix of archaic and modern traits that defy clear categorization into either group.
Research separated by nearly four decades uncovered fossils and artifacts from the Kibish Formation in the Lower Omo Valley in Ethiopia. These Omo Kibish hominins were represented by braincases and fragmented postcranial bones of three individuals found kilometers apart, dating back to around 233,000 years ago (Day 1969; McDougall, Brown, and Fleagle 2005; Vidal et al. 2022). One interesting finding was the variation in braincase size between the two more-complete specimens: while the individual named Omo I had a more globular dome, Omo II had an archaic-style long and low cranium.
Also in Ethiopia, a team led by Tim White (2003) excavated numerous fossils at Herto. There were fossilized crania of two adults and a child, along with fragments of more individuals. The dates ranged between 160,000 and 154,000 years ago. The skeletal traits and stone-tool assemblage were both intermediate between the archaic and modern types. Features reminiscent of modern humans included a tall braincase and thinner zygomatic (cheek) bones than those of archaic humans (Figure 13.7). Still, some archaic traits persisted in the Herto fossils, such as the supraorbital tori. Statistical analysis by other research teams concluded that at least some cranial measurements fit just within the modern human range (McCarthy and Lucas 2014), favoring categorization with our own species.

The timeline of material culture suggests a long period of relying on similar tools before a noticeable diversification of artifacts types. Researchers label the time of stable technology shared with archaic types the Middle Stone Age, while the subsequent time of diversification in material culture is called the Later Stone Age.
In the Middle Stone Age, the sites of Jebel Irhoud, Omo, and Herto all bore tools of the same flaked style as archaic assemblages, even though they were separated by almost 150,000 years. The consistency in technology may be evidence that behavioral modernity was not so developed. No clear signs of art dating back this far have been found either. Other hypotheses not related to behavioral modernity could explain these observations. The tool set may have been suitable for thriving in Africa without further innovation. Maybe works of art from that time were made with media that deteriorated or perhaps such art was removed by later humans.
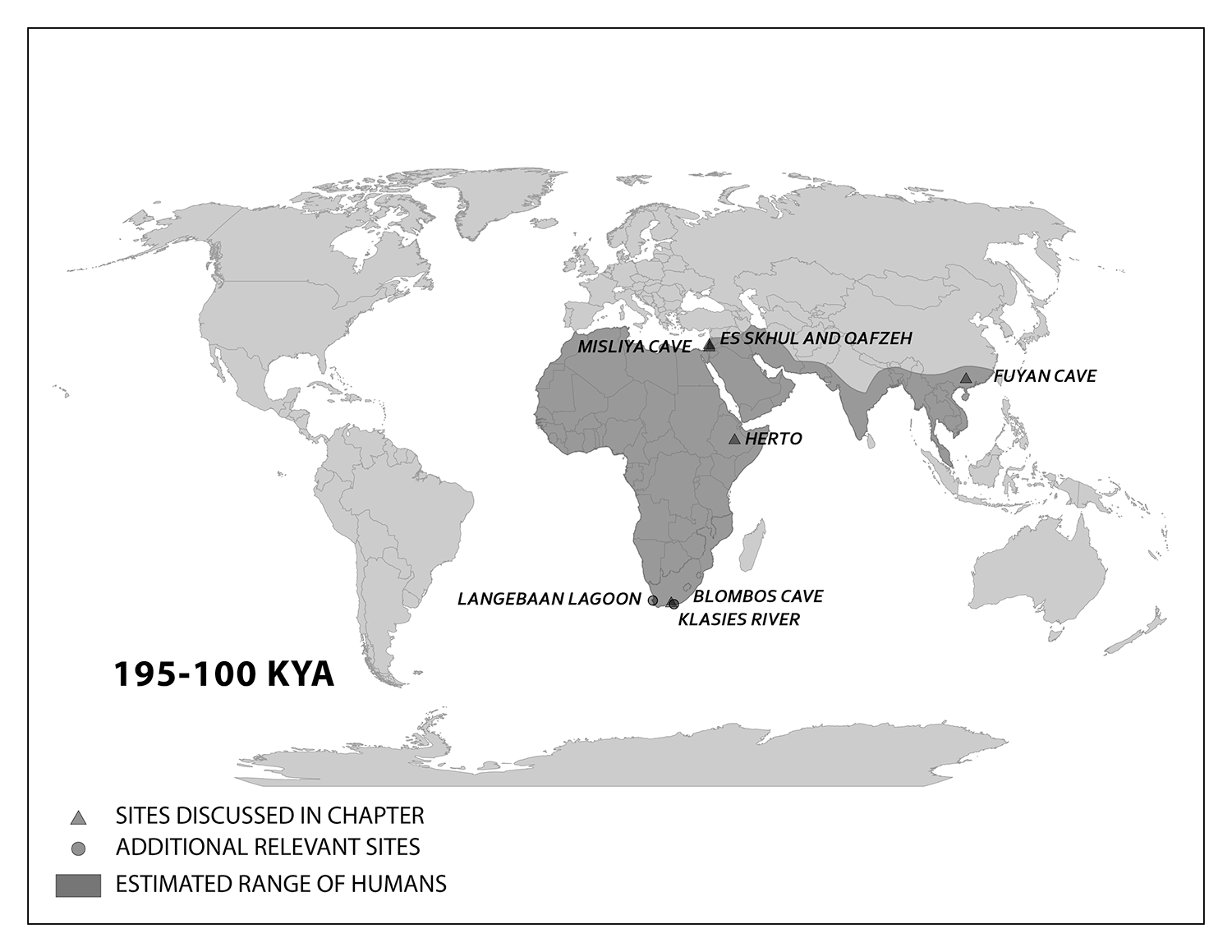
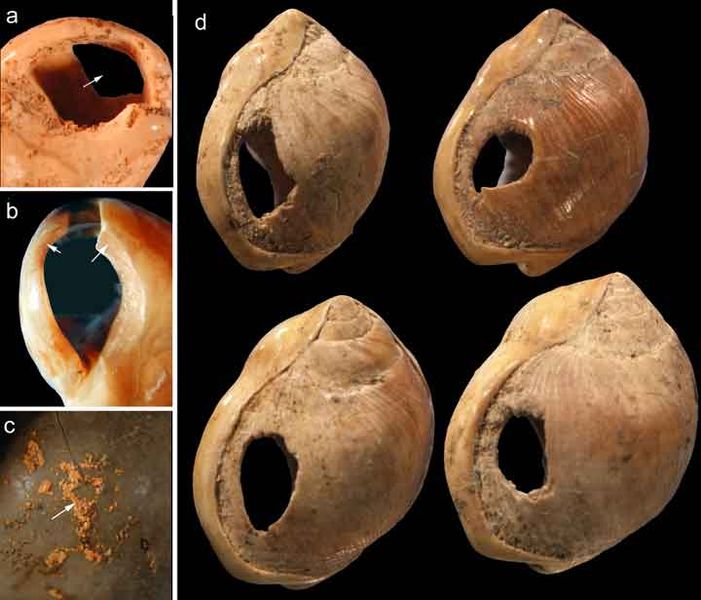
Evidence of what Homo sapiens did in Africa from the end of the Middle Stone Age to the Later Stone Age is concentrated in South African cave sites that reveal the complexity of human behavior at the time. For example, Blombos Cave, located along the present shore of the Cape of Africa facing the Indian Ocean, is notable for having a wide variety of artifacts. The material culture shows that toolmaking and artistry were more complex than previously thought for the Middle Stone Age. In a layer dated to 100,000 years ago, researchers found two intact ochre-processing kits made of abalone shells and grinding stones (Henshilwood et al. 2011). Marine snail shell beads from 75,000 years ago were also excavated (Figure 13.8; d’Errico et al. 2005). Together, the evidence shows that the Middle Stone Age occupation at Blombos Cave incorporated resources from a variety of local environments into their culture, from caves (ochre), open land (animal bones and fat), and the sea (abalone and snail shells). This complexity shows a deep knowledge of the region’s resources and their use—not just for survival but also for symbolic purposes.
On the eastern coast of South Africa, Border Cave shows new African cultural developments at the start of the Later Stone Age. Paola Villa and colleagues (2012) identified several changes in technology around 43,000 years ago. Stone-tool production transitioned from a slower process to one that was faster and made many microliths, small and precise stone tools. Changes in decorations were also found across the Later Stone Age transition. Beads were made from a new resource: fragments of ostrich eggs shaped into circular forms resembling present-day breakfast cereal O’s (d’Errico et al. 2012). These beads show a higher level of altering one’s own surroundings and a move from the natural to the abstract in terms of design.
Expansion into the Middle East and Asia
While modern Homo sapiens lived across Africa, some members eventually left the continent. These pioneers could have used two connections to the Middle East or West Asia. From North Africa, they could have crossed the Sinai Peninsula and moved north to the Levant, or eastern Mediterranean. Finds in that region show an early modern human presence. Other finds support the Southern Dispersal model, with a crossing from East Africa to the southern Arabian Peninsula through the Straits of Bab-el-Mandeb. It is tempting to think of one momentous event in which people stepped off Africa and into the Middle East, never to look back. In reality, there were likely multiple waves of movement producing gene flow back and forth across these regions as the overall range pushed east. The expanding modern human population could have thrived by using resources along the southern coast of the Arabian Peninsula to South Asia, with side routes moving north along rivers. The maximum range of the species then grew across Asia.
Modern Homo sapiens in the Middle East
Geographically, the Middle East is the ideal place for the African modern Homo sapiens population to inhabit upon expanding out of their home continent. In the Eastern Mediterranean coast of the Levant, there is a wealth of skeletal and material culture linked to modern Homo sapiens. Recent discoveries from Saudi Arabia further add to our view of human life just beyond Africa.
The Caves of Mount Carmel in present-day Israel have preserved skeletal remains and artifacts of modern Homo sapiens, the first-known group living outside Africa. The skeletal presence at Misliya Cave is represented by just part of the left upper jaw of one individual, but it is notable for being dated to a very early time, between 194,000 and 177,000 years ago (Hershkovitz et al. 2018). Later, from 120,000 to 90,000 years ago, fossils of multiple individuals across life stages were found in the caves of Es-Skhul and Qafzeh (Shea and Bar-Yosef 2005). The skeletons had many modern Homo sapiens traits, such as globular crania and more gracile postcranial bones when compared to Neanderthals. Still, there were some archaic traits. For example, the adult male Skhul V also possessed what researchers Daniel Lieberman, Osbjorn Pearson, and Kenneth Mowbray (2000) called marked or clear occipital bunning. Also, compared to later modern humans, the Mount Carmel people were more robust. Skhul V had a particularly impressive brow ridge that was short in height but sharply jutted forward above the eyes (Figure 13.9). The high level of preservation is due to the intentional burial of some of these people. Besides skeletal material, there are signs of artistic or symbolic behavior. For example, the adult male Skhul V had a boar’s jaw on his chest. Similarly, Qafzeh 11, a juvenile with healed cranial trauma, had an impressive deer antler rack placed over his torso (Figure 13.10; Coqueugniot et al. 2014). Perforated seashells colored with ochre, mineral-based pigment, were also found in Qafzeh (Bar-Yosef Mayer, Vandermeersch, and Bar-Yosef 2009).
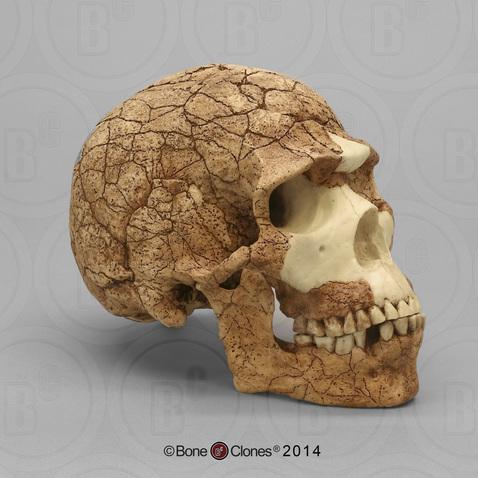

One remaining question is, what happened to the modern humans of the Levant after 90,000 years ago? Another site attributed to our species did not appear in the region until 47,000 years ago. Competition with Neanderthals may have accounted for the disappearance of modern human occupation since the Neanderthal presence in the Levant lasted longer than the dates of the early modern Homo sapiens. John Shea and Ofer Bar-Yosef (2005) hypothesized that the Mount Carmel modern humans were an initial expansion from Africa that failed. Perhaps they could not succeed due to competition with the Neanderthals who had been there longer and had both cultural and biological adaptations to that environment.
Modern Homo sapiens of China
A long history of paleoanthropology in China has found ample evidence of modern human presence. Four notable sites are the caves at Fuyan, Liujiang, Tianyuan, and Zhoukoudian. In the distant past, these caves would have been at least seasonal shelters that unintentionally preserved evidence of human presence for modern researchers to discover.
At Fuyan Cave in Southern China, paleoanthropologists found 47 adult teeth associated with cave formations dated to between 120,000 and 80,000 years ago (Liu et al. 2015). It is currently the oldest-known modern human site in China, though other researchers question the validity of the date range (Michel et al. 2016). The teeth have the small size and gracile features of modern Homo sapiens dentition.
The fossil Liujiang (or Liukiang) hominin (67,000 years ago) has derived traits that classified it as a modern Homo sapiens, though primitive archaic traits were also present. In the skull, which was found nearly complete, the Liujiang hominin had a taller forehead than archaic Homo sapiens but also had an enlarged occipital region (Figure 13.11; Brown 1999; Wu et al. 2008). Other parts of the skeleton also had a mix of modern and archaic traits: for example, the femur fragments suggested a slender length but with thick bone walls (Woo 1959).
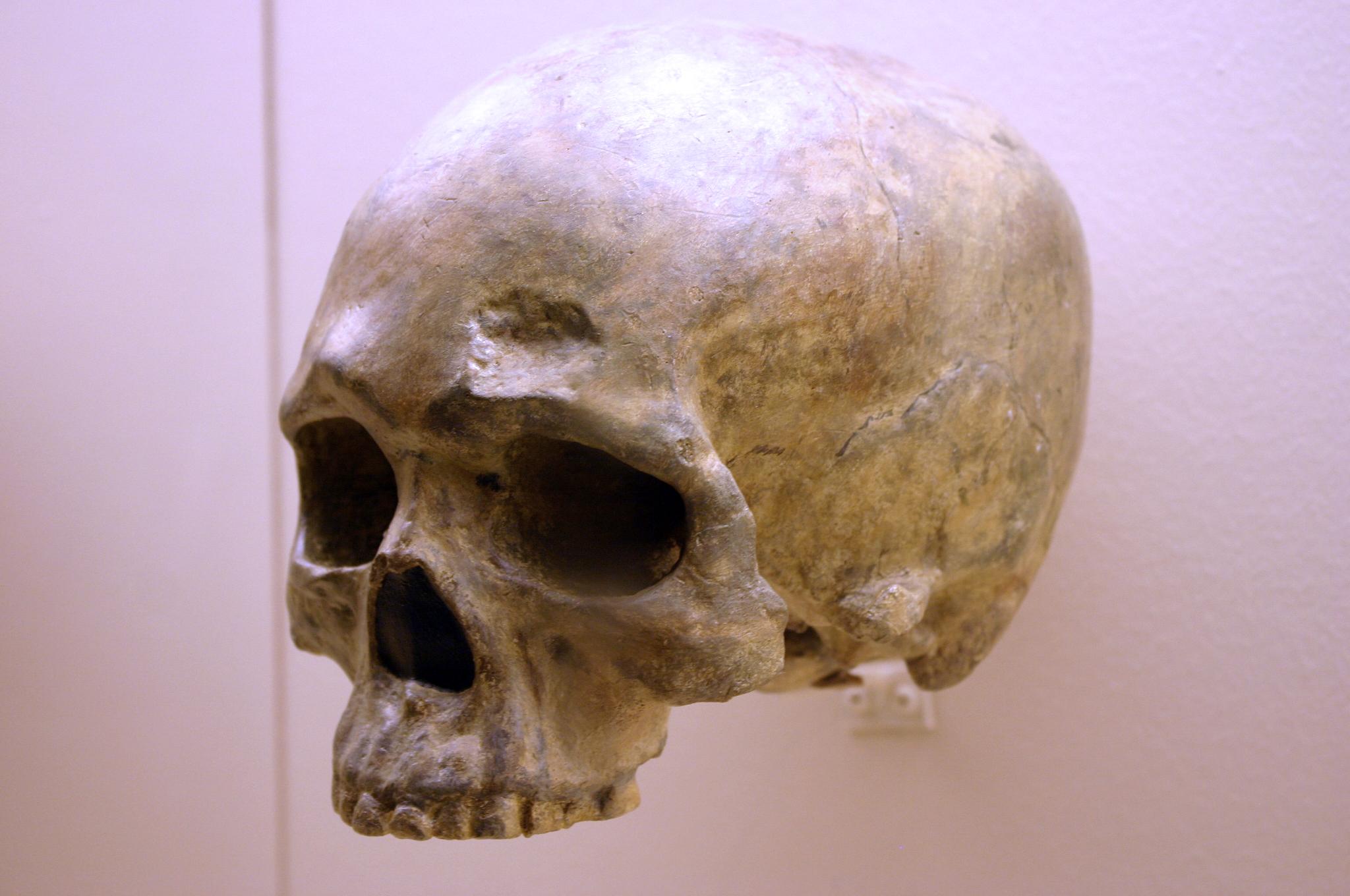
Another Chinese site to describe here is the one that has been studied the longest. In the Zhoukoudian Cave system (Figure 13.12), where Homo erectus and archaic Homo sapiens have also been found, there were three crania of modern Homo sapiens. These crania, which date to between 34,000 and 10,000 years ago, were all more globular than those of archaic humans but still lower and longer than those of later modern humans (Brown 1999; Harvati 2009). When compared to one another, the crania showed significant differences from one another. Comparison of cranial measurements to other populations past and present found no connection with modern East Asians, again showing that human variation was very different from what we see today.

Crossing to Australia
Expansion of the first modern human Asians, still following the coast, eventually entered an area that researchers call Sunda before continuing on to modern Australia. Sunda was a landmass made up of the modern-day Malay Peninsula, Sumatra, Java, and Borneo. Lowered sea levels connected these places with land bridges, making them easier to traverse. Proceeding past Sunda meant navigating Wallacea, the archipelago that includes the Indonesian islands east of Borneo. In the distant past, there were many megafauna, large animals that migrating humans would have used for food and materials (such as utilizing animals’ hides and bones). Further southeast was another landmass called Sahul, which included New Guinea and Australia as one contiguous continent. Based on fossil evidence, this land had never seen hominins or any other primates before modern Homo sapiens arrived. Sites along this path offer clues about how our species handled the new environment to live successfully as foragers.

The skeletal remains at Lake Mungo, land traditionally owned by Mutthi Mutthi, Ngiampaa, and Paakantji peoples, are the oldest known in the continent. The now-dry lake was one of a series located along the southern coast of Australia in New South Wales, far from where the first people entered from the north (Barbetti and Allen 1972; Bowler et al. 1970). Two individuals dating to around 40,000 years ago show signs of artistic and symbolic behavior, including intentional burial. The bones of Lake Mungo 1 (LM1), an adult female, were crushed repeatedly, colored with red ochre, and cremated (Bowler et al. 1970). Lake Mungo 3 (LM3), a tall, older male with a gracile cranium but robust postcranial bones, had his fingers interlocked over his pelvic region (Brown 2000).
Kow Swamp, within traditional Yorta Yorta land also in southern Australia, contained human crania that looked distinctly different from the ones at Lake Mungo (Durband 2014; Thorne and Macumber 1972). The crania, dated between 9,000 and 20,000 years ago, had extremely robust brow ridges and thick bone walls, but these were paired with globular features on the braincase (Figure 13.13).
While no fossil humans have been found at the Madjedbebe rock shelter in the North Territory of Australia, more than 10,000 artifacts found there show both behavioral modernity and variability (Clarkson et al. 2017). They include a diverse array of stone tools and different shades of ochre for rock art, including mica-based reflective pigment (similar to glitter). These impressive artifacts are as far back as 56,000 years old, providing the date for the earliest-known presence of humans in Australia.
From the Levant to Europe
The first modern human expansion into Europe occurred after other members of our species settled in East Asia and Australia. As the evidence from the Levant suggests, modern human movement to Europe may have been hampered by the presence of Neanderthals. It is suggested that another obstacle was the colder climate, which was incompatible with the biology of modern Homo sapiens from Africa, as they were adapted to high temperatures and ultraviolet radiation. Still, by 40,000 years ago, modern Homo sapiens had a detectable presence. This time was also the start of the Later Stone Age or Upper Paleolithic, when there was an expansion in cultural complexity. There is a wealth of evidence from this region due to a Western bias in research, the proximity of these findings to Western scientific institutions, and the desire of Western scientists to explore their own past.
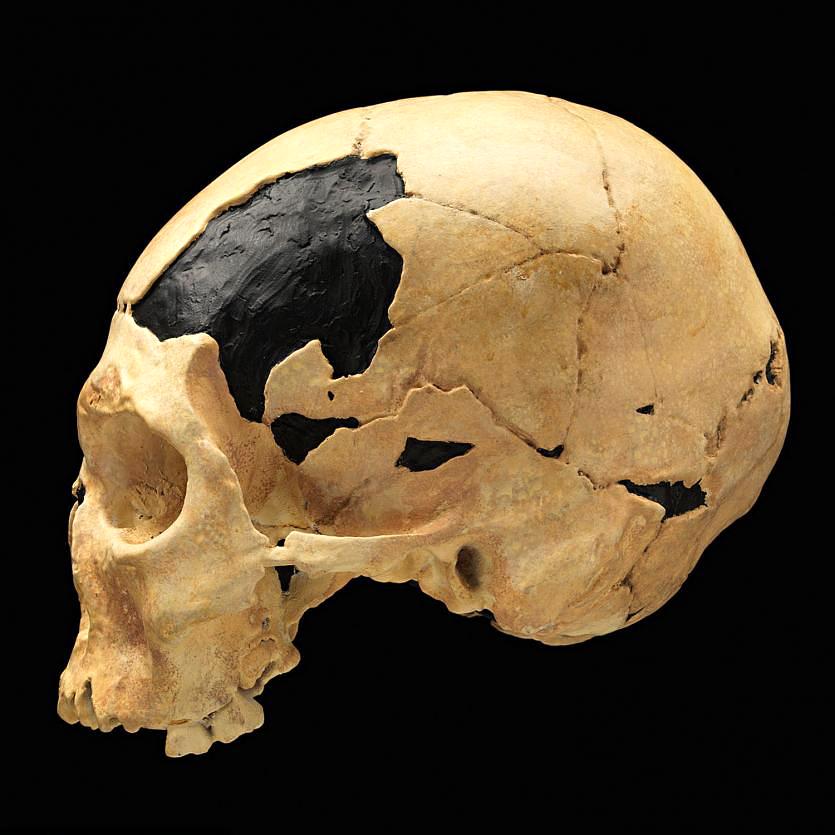
In Romania, the site of Peștera cu Oase (Cave of Bones) had the oldest-known remains of modern Homo sapiens in Europe, dated to around 40,000 years ago (Trinkaus et al. 2003a). Among the bones and teeth of many animals were the fragmented cranium of one person and the mandible of another (the two bones did not fit each other). Both bones have modern human traits similar to the fossils from the Middle East, but they also had Neanderthal traits. Oase 1, the mandible, had a mental eminence but also extremely large molars (Trinkaus et al. 2003b). This mandible has yielded DNA that surprisingly is equally similar to DNA from present-day Europeans and Asians (Fu et al. 2015). This means that Oase 1 was not the direct ancestor of modern Europeans. The Oase 2 cranium has the derived traits of reduced brow ridges along with archaic wide zygomatic cheekbones and an occipital bun (Figure 13.14; Rougier et al. 2007).
Dating to around 26,000 years ago, Předmostí near Přerov in the Czech Republic was a site where people buried over 30 individuals along with many artifacts. Eighteen individuals were found in one mass burial area, a few covered by the scapulae of woolly mammoths (Germonpré, Lázničková-Galetová, and Sablin 2012). The Předmostí crania were more globular than those of archaic humans but tended to be longer and lower than in later modern humans (Figure 13.15; Velemínská et al. 2008). The height of the face was in line with modern residents of Central Europe. There was also skeletal evidence of dog domestication, such as the presence of dog skulls with shorter snouts than in wild wolves (Germonpré, Lázničková-Galetová, and Sablin et al. 2012). In total, Předmostí could have been a settlement dependent on mammoths for subsistence and the artificial selection of early domesticated dogs.
The sequence of modern Homo sapiens technological change in the Later Stone Age has been thoroughly dated and labeled by researchers working in Europe. Among them, the Gravettian tradition of 33,000 years to 21,000 years ago is associated with most of the known curvy female figurines, often assumed to be “Venus” figures. Hunting technology also advanced in this time with the first known boomerang, atlatl (spear thrower), and archery. The Magdalenian tradition spread from 17,000 to 12,000 years ago. This culture further expanded on fine bone tool work, including barbed spearheads and fishhooks (Figure 13.16).
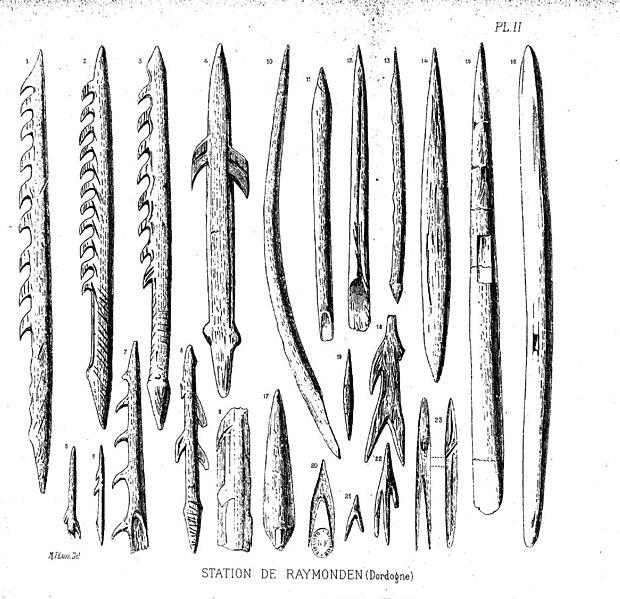
Among the many European sites dating to the Later Stone Age, the famous cave art sites deserve mention. Chauvet-Pont-d’Arc Cave in southern France dates to separate Aurignacian occupations 31,000 years ago and 26,000 years ago. Over a hundred art pieces representing 13 animal species are preserved, from commonly depicted deer and horses to rarer rhinos and owls. Another French cave with art is Lascaux, which is several thousand years younger at 17,000 years ago in the Magdalenian period. At this site, there are over 6,000 painted figures on the walls and ceiling (Figure 13.17). Scaffolding and lighting must have been used to make the paintings on the walls and ceiling deep in the cave. Overall, visiting Lascaux as a contemporary must have been an awesome experience: trekking deeper in the cave lit only by torches giving glimpses of animals all around as mysterious sounds echoed through the galleries.

Special Topic: Cannibalism and Culture – Mortuary Practices in Modern Homo sapiens
Within a 2017 publication in the Journal of Archaeological Method and Theory, Saladié and Rodríguez-Hidalgo bring light to traces of early cannibalism in western Eurasia, arguing that context-specific cannibalistic practices were present throughout the Pleistocene and increased notably from the end of the Upper Palaeolithic and onward (Saladié & Rodriguez-Hidalgo, 2017). While early hominins and Neandertals are recognized in this research, the authors highlight the presence of these mortuary practices in a cluster of Homo sapiens sites. More recent research uncovers similar findings that back these claims as well, where human bones in Herto Ethiopia, Maszycka Cave Poland, and Gough’s Cave in the United Kingdom show anthropogenic defleshing and other modifications which have been interpreted as cannibalism (Pobiner, Et al. 2023). These findings suggest that cannibalistic behaviours formed a recurring aspect of modern human behaviour in certain ecological and cultural contexts.

A significant example comes from the Neolithic levels of Fontbrégua Cave in southeastern France, where Paola Villa and colleagues compared clusters of human bones with the remains of wild and domestic animals from the same sediments. The study found that human bodies were butchered, processed, and most likely eaten in a way that parallels animal carcass treatment, including the placement and timing of cut marks, dismemberment sequences, and perimortem fractures to open marrow cavities (Villa, Et al. 1986). As the assemblage comes from a primary depositional context with pristine preservation and careful excavation, the authors suggest that cannibalism is the only satisfactory explanation for the pattern of cut marks and breakage seen on the human bones.
More recent work has furthered this hypothesis for Late Upper Palaeolithic Homo sapiens, especially in Magdalenian contexts. In a 2023 study, researchers combined archeological and genetic evidence from fifty-nine Magdalenian sites, concluding that this specific culture shows an unusually high frequency of cannibalistic cases compared to earlier and later hominin groups; so much so that they identify “primary burial and cannibalism” as the two main mortuary expressions (Marsh & Bello, 2023). Additionally, new analyses from Maszycka Cave in Poland have described cut and broken human bones in patterns consistent with human consumption, reinforcing this cannibalism hypothesis (Marginedas & Saladié, 2025). Together, these studies suggest that for some Magdalenian groups, mortuary cannibalism was a habitual way of disposing of the dead, not just a one-off crisis response. At the same time, researchers emphasize that not every modified skeleton indicates blatant consumption of the dead and that ritual or symbolic perspectives must also be considered. Previously noted in chapter eleven, Ullrich’s survey of European mortuary practices presents frequent manipulations of corpses from the Palaeolithic through the Hallstatt period. Said manipulations include cut marks, dismemberment, skull fracturing, and marrow extraction (Ullrich, 2005, p. 258), which Ullrich interprets as cannibalistic rites embedded in cult ceremonies rather than everyday subsistence. In the author’s view, some Palaeolithic groups may have believed that consuming members of their community allowed them to take on the strengths,

abilities, or even mental aspects of their dead member; the act of cannibalism may have functioned as a form of appropriating physical and mental powers rather than simply obtaining calories. With that, Neolithic assemblages show how careful taphonomic work can distinguish cuts linked to interpersonal violence from those associated with systemic butchery (Marginedas & Saladié, 2025). The findings seek to highlight that some Homo sapiens populations combined ritual, mortuary, and nutritional motives when processing human remains.
These past and contemporary findings are significant for future anthropology students as they shed light on biological anthropology methodology and interpretation. Archaeologists are able to distinguish between occasions when humans were handled like any other carcass and times when they were handled in more symbolic ways thanks to factors like cut mark orientation, the timing of bone breakage, and direct comparison with animal remains. Readers interested in exploring this topic further should investigate the context-specific motivations for cannibalism, like nutritional stress, warfare, funerary sites, or ritual power appropriation.
Similar archeological techniques have been used to explore behaviours of cannibalism among Indigenous Huron-Wendat populations in 1651 Canada, where human bones exhibit cutmarks, perimortem fractures, and thermal modifications (Spence & Jackson, 2014). These shared taphonomic signatures across continents reveal recurring practices under ecological stress, bridging prehistoric Europe to North American contexts. Engaging with such analytical techniques opens up new ways that archeologists and anthropologists can investigate the variability in human behaviour, from nutritional crises to mortuary rituals.
Peopling of the Americas
By 25,000 years ago, our species was the only member of Homo left on Earth. Gone were the Neanderthals, Denisovans, Homo naledi, and Homo floresiensis. The range of modern Homo sapiens kept expanding eastward into—using the name given to this area by Europeans much later—the Western Hemisphere. This section will address what we know about the peopling of the Americas, from the first entry to these continents to the rapid spread of Indigenous Americans across its varied environments.
While evidence points to an ancient land bridge called Beringia that allowed people to cross from what is now northeastern Siberia into modern-day Alaska, what people did to cross this land bridge is still being investigated. For most of the 20th century, the accepted theory was the Ice-Free Corridor model. It stated that northeast Asians (East Asians and Siberians) first expanded across Beringia inland through a passage between glaciers that opened into the western Great Plains of the United States, just east of the Rocky Mountains, around 13,000 years ago (Swisher et al. 2013). While life up north in the cold environment would have been harsh, migrating birds and an emerging forest might have provided sustenance as generations expanded through this land (Potter et al. 2018).
However, in recent decades, researchers have accumulated evidence against the Ice-Free Corridor model. Archaeologist K. R. Fladmark (1979) brought the alternate Coastal Route model into the archaeological spotlight; researcher Jon M. Erlandson has been at the forefront of compiling support for this theory (Erlandson et al. 2015). The new focus is the southern edge of the land bridge instead of its center: About 16,000 years ago, members of our species expanded along the coastline from northeast Asia, east through Beringia, and south down the Pacific Coast of North America while the inland was still sealed off by ice. The coast would have been free of ice at least part of the year, and many resources would have been found there, such as fish (e.g., salmon), mammals (e.g., whales, seals, and otters), and plants (e.g., seaweed).
South through the Americas
When the first modern Homo sapiens reached the Western Hemisphere, the spread through the Americas was rapid. Multiple migration waves crossed from North to South America (Posth et al. 2018). Our species took advantage of the lack of hominin competition and the bountiful resources both along the coasts and inland. The Americas had their own wide array of megafauna, which included woolly mammoths (Figure 13.20), mastodons, camels, horses, ground sloths, giant tortoises, and—a favorite of researchers—a two-meter-tall beaver. The reason we cannot see these amazing animals today may be that resources gained from these fauna were crucial to the survival for people over 12,000 years ago (Araujo et al. 2017). Several sites are notable for what they add to our understanding of the distant past in the Americas, including interactions with megafauna and other elements of the environment.

A 2019 discovery may allow researchers to improve theories about the peopling of the Americas. In White Sands National Park, New Mexico, 60 human footprints have been astonishingly dated to around 22,000 years ago (Bennett et al. 2021). This date and location do not match either the Ice-Free Corridor or Coastal Route models. Researchers are now working to verify the find and adjust previous models to account for the new evidence. This groundbreaking find is sparking new theories; it is another example of the fast pace of research performed on our past.
Monte Verde is a landmark site that shows that the human population had expanded down the whole vertical stretch of the Americas to Chile by 14,600 years ago. The site has been excavated by archaeologist Tom D. Dillehay and his team (2015). The remains of nine distinct edible species of seaweed at the site shows familiarity with coastal resources and relates to the Coastal Route model by showing a connection between the inland people and the sea.

Named after the town in New Mexico, the Clovis stone-tool style is the first example of a widespread culture across much of North America, between 13,400 and 12,700 years ago (Miller, Holliday, and Bright 2013). Clovis points were fluted with two small projections, one on each end of the base, facing away from the head (Figure 13.21). The stone points found at this site match those found as far as the Canadian border and northern Mexico, and from the west coast to the east coast of the United States. Fourteen Clovis sites also contained the remains of mammoths or mastodons, suggesting that hunting megafauna with these points was an important part of life for the Clovis people. After the spread of the Clovis style, it diversified into several regional styles, keeping some of the Clovis form but also developing their own unique touches.
The Big Picture: The Assimilation Hypothesis
How do researchers make sense of all of these modern Homo sapiens discoveries that cover over 300,000 years of time and stretch across every continent except Antarctica? How was modern Homo sapiens related to archaic Homo sapiens?
The Assimilation hypothesis proposes that modern Homo sapiens evolved in Africa first and expanded out but also interbred with the archaic Homo sapiens they encountered outside Africa (Figure 13.22). This hypothesis is powerful since it explains why Africa has the oldest modern human fossils, why early modern humans found in Europe and Asia bear a resemblance to the regional archaics, and why traces of archaic DNA can be found in our genomes today (Dannemann and Racimo 2018; Reich et al. 2010; Reich et al. 2011; Slatkin and Racimo 2016; Smith et al. 2017; Wall and Yoshihara Caldeira Brandt 2016).
While researchers have produced a model that satisfies the data, there are still a lot of questions for paleoanthropologists to answer regarding our origins. What were the patterns of migration in each part of the world? Why did the archaic humans go extinct? In what ways did archaic and modern humans interact? The definitive explanation of how our species started and what our ancestors did is still out there to be found. You are now in a great place to welcome the next discovery about our distant past—maybe you’ll even contribute to our understanding as well.
The Chain Reaction of Agriculture
While it may be hard to imagine today, for most of our species’ existence we were nomadic: moving through the landscape without a singular home. Instead of a refrigerator or pantry stocked with food, we procured nutrition and other resources as needed based on what was available in the environment. This section gives an overview of how the foraging lifestyle enabled the expansion of our species and how the invention of a new way of life caused a chain reaction of cultural change.
The Foraging Tradition
There are a variety of possible subsistence strategies, or methods of finding sustenance and resources. To understand our species is to understand the subsistence strategy of foraging, or the search for resources in the environment. While most (but not all) humans today live in cultures that practice agriculture (whereby we greatly shape the environment to mass produce what we need), we have spent far more time as nomadic foragers than as settled agriculturalists. As such, it has been suggested that our traits have evolved to be primarily geared toward foraging. For instance, our efficient bipedalism allows persistence-hunting across long distances as well as movement from resource to resource.
How does human foraging, also known as hunting and gathering, work? Anthropologists have used all four fields to answer this question (see Ember n.d.). Typically, people formed bands, or kin-based groups of around 50 people or less (rarely over 100). A band’s organization would be egalitarian, with a flexible hierarchy based on an individual’s age, level of experience, and relationship with others. Everyone would have a general knowledge of the skills assigned to their gender roles, rather than specializing in different occupations. A band would be able to move from place to place in the environment, using knowledge of the area to forage (Figure 13.23). In varied environments—from savannas to tropical forests, deserts, coasts, and the Arctic circle—people found sustenance needed for survival.

Humans made extensive use of the foraging subsistence strategy, but this lifestyle did have limitations. The ease of foraging depended on the richness of the environment. Due to the lack of storage, resources had to be dependably found when needed. While a bountiful environment would require just a few hours of foraging a day and could lead to a focus on one location, the level and duration of labor increased greatly in poor or unreliable environments. Labor was also needed to process the acquired resources, which contributed to the foragers’ daily schedule (Crittenden and Schnorr 2017).
The adaptations to foraging found in modern Homo sapiens may explain why our species became so successful both within Africa and in the rapid expansion around the world. Overcoming the limitations, each generation at the edge of our species’s range would have found it beneficial to expand a little further, keeping contact with other bands but moving into unexplored territory where resources were more plentiful. The cumulative effect would have been the spread of modern Homo sapiens across continents and hemispheres.
Why Agriculture?
After hundreds of thousands of years of foraging, some groups of people around 12,000 years ago started to practice agriculture. This transition, called the Neolithic Revolution, occurred at the start of the Holocene epoch. While the reasons for this global change are still being investigated, two likely co-occurring causes are a growing human population and natural global climate change.
Overcrowding could have affected the success of foraging in the environment, leading to the development of a more productive subsistence strategy (Cohen 1977). Foraging works best with low population densities since each band needs a lot of space to support itself. If too many people occupy the same environment, they deplete the area faster. The high population could exceed the carrying capacity, or number of people a location can reliably support. Reaching carrying capacity on a global level due to growing population and limited areas of expansion would have been an increasingly pressing issue after the expansion through the major continents by 14,600 years ago.
A changing global climate immediately preceded the transition to agriculture, so researchers have also explored a connection between the two events. Since the Last Glacial Maximum of 23,000 years ago, the Earth slowly warmed. Then, from 13,000 to 11,700 years ago, the temperature in most of the Northern Hemisphere dropped suddenly in a phenomenon called the Younger Dryas. Glaciers returned in Europe, Asia, and North America. In Mesopotamia, which includes the Levant, the climate changed from warm and humid to cool and dry. The change would have occurred over decades, disrupting the usual nomadic patterns and subsistence of foragers around the world. The disruption to foragers due to the temperature shift could have been a factor in spurring a transition to agriculture. Researchers Gregory K. Dow and colleagues (2009) believe that foraging bands would have clustered in the new resource-rich places where people started to direct their labor to farming the limited area. After the Younger Dryas ended, people expanded out of the clusters with their agricultural knowledge (Figure 13.24).
The double threat of the limitation of human continental expansion and the sudden global climate change may have placed bands in peril as more populations outpaced their environment’s carrying capacity. Not only had a growing population led to increased competition with other bands, but environments worldwide had shifted to create more uncertainty. As such, it has been proposed that as people in different areas around the world faced this unpredictable situation, they became the independent inventors of agriculture.
Agriculture around the World
Due to global changes to the human experience starting from 12,000 years ago, it has been suggested that cultures with no knowledge of each other turned toward intensely farming their local resources (see Figure 13.24). It is proposed that the first farmers engaged in artificial selection of their domesticates to enhance useful traits over generations. The switch to agriculture took time and effort with no guarantee of success and constant challenges (e.g. fires, droughts, diseases, and pests). The regions with the most widespread impact in the face of these obstacles became the primary centers of agriculture (Figure 13.25; Fuller 2010):
- Mesopotamia: The Fertile Crescent from the Tigris and Euphrates rivers through the Levant was where bands started to domesticate plants and animals around 12,000 years ago. The connection between the development of agriculture and the Younger Dryas was especially strong here. Farmed crops included wheat, barley, peas, and lentils. This was also where cattle, pigs, sheep, and goats were domesticated.
- South and East Asia: Multiple regions across this land had varieties of rice, millet, and soybeans by 10,000 years ago. Pigs were farmed with no connection to Mesopotamia. Chickens were also originally from this region, bred for fighting first and food second.
- New Guinea: Agriculture started here 10,000 years ago. Bananas, sugarcane, and taro were native to this island. Sweet potatoes were brought back from voyages to South America around the year C.E. 1000. No known animal farming occurred here.
- Mesoamerica: Agriculture from Central Mexico to northern South America also occurred from 10,000 years ago; it was also only plant based. Maize was a crop bred from teosinte grass, which has become one of the global staples. Beans, squash, and avocados were also grown in this region.
- The Andes: Starting around 8,000 years ago, local domesticated plants started with squash but later included potatoes, tomatoes, beans, and quinoa. Maize was brought down from Mesoamerica. The main farm animals were llamas, alpacas, and guinea pigs.
- Sub-Saharan Africa: This region went through a change 5,000 years ago called the Bantu expansion. The Bantu agriculturalists were established in West Central Africa and then expanded south and east. Native varieties of rice, yams, millet, and sorghum were grown across this area. Cattle were also domesticated here.
- Eastern North America: This region was the last major independent agriculture center, from 4,000 years ago. Squash and sunflower are the produce from this region that are most known today, though sumpweed and pitseed goosefoot were also farmed. Hunting was still the main source of animal products.

By 5,000 years ago, our species was well within the Neolithic Revolution. Agriculturalists spread to neighboring parts of the world with their domesticates, further expanding the use of this subsistence strategy. From this point, the human species changed from being primarily foragers to primarily agriculturalists with skilled control of their environments. The planet changed from mostly unaffected by human presence to being greatly transformed by humans. The revolution took millennia, but it was a true revolution as our species’ lifestyle was dramatically reshaped.
Cultural Effects of Agriculture
The worldwide adoption of agriculture altered the course of human culture and history forever. The core change in human culture due to agriculture is the move toward not moving: rather than live a nomadic lifestyle, farmers had to remain in one area to tend to their crops and livestock. The term for living bound to a certain location is sedentarism. This led to new aspects of life that were uncommon among foragers: the construction of permanent shelters and agricultural infrastructure, such as fields and irrigation, plus the development of storage technology, such as pottery, to preserve extra resources in case of future instability.

The high productivity of successful agriculture sparked further changes (Smith 2009). It is argued that since successful agriculture produced a much greater amount of food and other resources per unit of land compared to foraging, the population growth rate skyrocketed. The surplus of a bountiful harvest also provided insurance for harder times, reducing the risk of famine. Changes happened to society as well. With a few farming households producing enough food to feed many others, other people could focus on other tasks. So began specialization into different occupations such as craftspeople, traders, religious figures, and artists, spurring innovation in these areas as people could now devote time and effort toward specific skills. These interdependent people would settle an area together for convenience. The growth of these settlements led to urbanization, the founding of cities that became the foci of human interaction (Figure 13.26).
The formation of cities led to new issues that sparked the growth of further specializations, called institutions. These are cultural constructs that exist beyond the individual and have wide control over a population. Leadership of these cities became hierarchical with different levels of rank and control. The stratification of society increased social inequality between those with more or less power over others. Under leadership, people built impressive monumental architecture, such as pyramids and palaces, that embodied the wealth and power of these early cities. Alliances could unite cities, forming the earliest states. In several regions of the world, state organization expanded into empires, wide-ranging political entities that covered a variety of cultures.
Urbanization brought new challenges as well. The concentration of sedentary peoples was ideal for infectious diseases to thrive since they could jump from person to person and even from livestock to person (Armelagos, Brown, and Turner 2005). While successful agriculture provided a large surplus of food to thwart famine, the food produced offered less diverse food sources than foragers’ diets (Cohen and Armelagos 1984; Cohen and Crane-Kramer 2007). This shift in nutrition caused other diseases to flourish among those who adopted farming, such as dental cavities and malocclusion (the misalignment of teeth caused by soft, agricultural diets). The need to extract “wisdom teeth” or third molars seen in agricultural cultures today stems from this misalignment between the environment our ancestors adapted to and our lifestyles today.
As the new disease trends show, the adoption of agriculture and the ensuing cultural changes were not entirely positive. It is also important to note that this is not an absolutely linear progression of human culture from simple to complex. In many cases, empires have collapsed and, in some cases, cities dispersed to low-density bands that rejected institutions. However, a global trend has emerged since the adoption of agriculture, wherein population and social inequality have increased, leading to the massive and influential nation-states of today.
The rise of states in Europe has a direct impact on many of this book’s topics. Science started as a European cultural practice by the upper class that became a standardized way to study the world. Education became an institution to provide a standardized path toward producing and gaining knowledge. The scientific study of human diversity, embroiled in the race concept that still haunts us today, was connected to the European slave trade and colonialism.
Also starting in Europe, the Industrial Revolution of the 19th century turned cities into centers of mass manufacturing and spurred the rapid development of inventions (Figure 13.27). In the technologically interconnected world of today, human society has reached a new level of complexity with globalization. In this system, goods are mass-produced and consumed in different parts of the world, weakening the reliance on local farms and factories. The imbalanced relationship between consumers and producers of goods further increases economic inequality.

As states based on agriculture and industry keep exerting influence on humanity today, there are people, like the Hadzabe of Tanzania, who continue to live a lifestyle centered on foraging. Due to the overwhelming force that agricultural societies exert, foragers today have been marginalized to live in the least habitable parts of the world—the areas that are not conducive to farming, such as tropical rainforests, deserts, and the Arctic (Headland et al. 1989). Foragers can no longer live in the abundant environments that humans would have enjoyed before the Neolithic Revolution. Interactions with agriculturalists are typically imbalanced, with trade and other exchanges heavily favoring the larger group. One of anthropology’s important roles today is to intelligently and humanely manage equitable interactions between people of different backgrounds and levels of influence.
Special Topic: Indigenous Land Management
Insight into the lives of past modern humans has evolved as researchers revise previous theories and establish new connections with Indigenous knowledge holders.
The outdated view of foraging held that people lived off of the land without leaving an impact on the environment. Accompanying this idea was anthropologist Marshall Sahlins’s (1968) proposal that foragers were the “original affluent society” since they were meeting basic needs and achieving satisfaction with less work hours than agriculturalists and city-dwellers. This view countered an earlier idea that foragers were always on the brink of starvation. Sahlins’s theory took hold in the public eye as an attractive counterpoint to our busy contemporary lives in which we strive to meet our endless wants.
A fruitful type of study involving researchers collaborating with Indigenous experts has found that foragers did not just live off the land with minimal effort nor were they barely surviving in unchanging environments. Instead, they shaped the landscape to their needs using labor and strategies that were more subtle than what European colonizers and subsequent researchers were used to seeing. Research from two regions shows the latest developments in understanding Indigenous land management.
In British Columbia, Canada, the bridging of scientific and Indigenous perspectives has shown that the forests of the region are not untouched wilderness but, rather, have been crafted by Indigenous peoples thousands of years ago. Forest gardens adjacent to archaeological sites show higher plant diversity than unmanaged places even after 150 years (Armstrong et al. 2021). On the coast, 3,500-year-old archaeological sites are evidence of constructed clam gardens, according to Indigenous experts (Lepofsky et al. 2015). Another project, in consultation with Elders of the T’exelc (William Lakes First Nation) in British Columbia, introduced researchers to explanations of how forests were managed before the practice was disrupted by European colonialism (Copes-Gerbitz et al. 2021). Careful management of controlled fires reduced the density of the forest to favor plants such as raspberries and allow easier movement through the landscape.
Similarly, the study of landscapes in Australia, in consultation with Aboriginal Australians today, shows that areas previously considered wilderness by scientists were actually the result of controlling fauna and fires. The presence of grasslands with adjacent forests were purposely constructed to attract kangaroos for hunting (Gammage 2008). People also managed other animal and insect life, from emus to caterpillars. In Tasmania, a shift from productive grassland to wildfire-prone rainforest occurred after Aboriginal Australian land management was replaced by British colonial rule (Fletcher, Hall, and Alexander 2021). The site of Budj Bim of the Gunditjmara people has archaeological features of aquaculture, or the farming of fish, that date back 6,600 years (McNiven et al. 2012; McNiven et al. 2015). These examples show that Indigenous knowledge of how to manipulate the environment may be invaluable at the state level, such as by creating an Aboriginal ranger program to guide modern land management.
The Future of Humanity
A common question stemming from understanding human evolution is: What will the genetic and biological traits of our species be hundreds of thousands of years in the future? When faced with this question, people tend to think of directional selection. Maybe our braincases will be even larger, resembling the large-headed and small-bodied aliens of science fiction (Figure 13.28). Or, our hands could be specialized for interacting with our touch-based technology with less risk of repetitive injury. These ideas do not stand up to scrutiny. Since natural selection is based on adaptations that increase reproductive success, any directional change must be due to a higher rate of producing successful offspring compared to other alleles. Larger brains and more agile fingers would be convenient to possess, but they do not translate into an increase in the underlying allele frequencies.
Scientists are hesitant to professionally speculate on the unknowable, and we will never know what is in store for our species one thousand or one million years from now, but there are two trends in human evolution that may carry on into the future: increased genetic variation and a reduction in regional differences.
Rather than a directional change, genetic variation in our species could expand. Our technology can protect us from extreme environments and pathogens, even if our biological traits are not tuned to handle these stressors. The rapid pace of technological advancement means that biological adaptations will become less and less relevant to reproductive success, so nonbeneficial genetic traits will be more likely to remain in the gene pool. Biological anthropologist Jay T. Stock (2008) views environmental stress as needing to defeat two layers of protection before affecting our genetics. The first layer is our cultural adaptations. Our technology and knowledge can reduce pressure on one’s genotype to be “just right” to pass to the next generation. The second defense is our flexible physiology, such as our acclimatory responses. Only stressors not handled by these powerful responses would then cause natural selection on our alleles. These shields are already substantial, and cultural adaptations will only keep increasing in strength.
The increasing ability to travel far from one’s home region means that there will be a mixing of genetic variation on a global level in the future of our species. In recent centuries, gene flow of people around the world has increased, creating admixture in populations that had been separated for tens of thousands of years. For skin color, this means that populations all around the world could exhibit the whole range of skin colors, rather than the current pattern of decreasing melanin pigment farther from the equator. The same trend of intermixing would apply to all other traits, such as blood types. While our genetics will become more varied, the variation will be more intermixed instead of regionally isolated.
Our distant descendants will not likely be dextrous ultraintellectuals; more likely, they will be a highly variable and mobile species supported by novel cultural adaptations that make up for any inherited biological limitations. Technology may even enable the editing of DNA directly, changing these trends. With the uncertainty of our future, these are just the best-educated guesses for now. Our future is open and will be shaped little by little by the environment, our actions, and the actions of our descendants.
Summary
Modern Homo sapiens is the species that took the hominin lifestyle the furthest to become the only living member of that lineage. The largest factor that allowed us to persist while other hominins went extinct was likely our advanced ability to culturally adapt to a wide variety of environments. Our species, with its skeletal and behavioral traits, was well-suited to be generalist-specialists who successfully foraged across most of the world’s environments. The biological basis of this adaptation was our reorganized brain that facilitated innovation in cultural adaptations and intelligence for leveraging our social ties and finding ways to acquire resources from the environment. As the brain’s ability increased, it shaped the skull by reducing the evolutionary pressure to have large teeth and robust cranial bones to produce the modern Homo sapiens face.
Our ability to be generalist-specialists is seen in the geographical range that modern Homo sapiens covered in 300,000 years. In Africa, our species formed from multiregional gene flow that loosely connected archaic humans across the continent. People then expanded out to the rest of the continental Eurasia and even further to the Americas.
For most of our species’s existence, foraging was the general subsistence strategy within which people specialized to culturally adapt to their local environment. With omnivorousness and mobility, people found ways to extract and process resources, shaping the environment in return. When resource uncertainty hit the species, people around the world focused on agriculture to have a firmer control of sustenance. The new strategy shifted human history toward exponential growth and innovation, leading to our high dependence on cultural adaptations today.
While a cohesive image of our species has formed in recent years, there is still much to learn about our past. The work of many driven researchers shows that there are amazing new discoveries made all the time that refine our knowledge of human evolution. Technological innovations such as DNA analysis enable scientists to approach lingering questions from new angles. The answers we get allow us to ask even more insightful questions that will lead us to the next revelation. Like the pink limestone strata at Jebel Irhoud, previous effort has taken us so far and you are now ready to see what the next layer of discovery holds.
Hominin Species Summary
|
Hominin |
Modern Homo sapiens |
|
Dates |
315,000 years ago to present |
|
Region(s) |
Starting in Africa, then expanding around the world |
|
Famous discoveries |
Cro-Magnon individuals, discovered 1868 in Dordogne, France. Otzi the Ice Man, discovered 1991 in the Alps between Austria and Italy. Kennewick man, discovered 1996 in Washington state. |
|
Brain size |
1400 cc average |
|
Dentition |
Extremely small with short cusps. |
|
Cranial features |
An extremely globular brain case and gracile features throughout the cranium. The mandibular symphysis forms a chin at the anterior-most point. |
|
Postcranial features |
Gracile skeleton adapted for efficient bipedal locomotion at the expense of the muscular strength of most other large primates. |
|
Culture |
Extremely extensive and varied culture with many spoken and written languages. Art is ubiquitous. Technology is broad in complexity and impact on the environment. |
|
Other |
The only living hominin. Chimpanzees and bonobos are the closest living relatives. |
Review Questions
- What are the skeletal and behavioral traits that define modern Homo sapiens? What are the evolutionary explanations for its presence?
- What are some creative ways that researchers have learned about the past by studying fossils and artifacts?
- How do the discoveries mentioned in “First Africa, Then the World” fit the Assimilation model?
- What is foraging? What adaptations do we have for this subsistence strategy? Could you train to be a skilled forager?
- What are aspects of your life that come from dependence on agriculture and its cultural effects? Where did the ingredients of your favorite foods originate from?
Key Terms
African multiregionalism: The idea that modern Homo sapiens evolved as a complex web of small regional populations with sporadic gene flow among them.
Agriculture: The mass production of resources through farming and domestication.
Aquaculture: The farming of fish using techniques such as trapping, channels, and artificial ponds.
Assimilation hypothesis: Current theory of modern human origins stating that the species evolved first in Africa and interbred with archaic humans of Europe and Asia.
Atlatl: A handheld spear thrower that increased the force of thrown projectiles.
Band: A small group of people living together as foragers.
Beringia: Ancient landmass that connected Siberia and Alaska. The ancestors of Indigenous Americans would have crossed this area to reach the Americas.
Carrying capacity: The amount of organisms that an environment can reliably support.
Coastal Route model: Theory that the first Paleoindians crossed to the Americas by following the southern coast of Beringia.
Early Modern Homo sapiens, Early Anatomically Modern Human: Terms used to refer to transitional fossils between archaic and modern Homo sapiens that have a mosaic of traits. Humans like ourselves, who mostly lack archaic traits, are referred to as Late Modern Homo sapiens and simply Anatomically Modern Humans.
Egalitarian: Human organization without strict ranks. Foraging societies tend to be more egalitarian than those based on other subsistence strategies.
Foraging: Lifestyle consisting of frequent movement through the landscape and acquiring resources with minimal storage capacity.
Generalist-specialist niche: The ability to survive in a variety of environments by developing local expertise. Evolution toward this niche may have been what allowed modern Homo sapiens to expand past the geographical range of other human species.
Globalization: A recent increase in the interconnectedness and interdependence of people that is facilitated with long-distance networks.
Globular: Having a rounded appearance. Increased globularity of the braincase is a trait of modern Homo sapiens.
Gracile: Having a smooth and slender quality; the opposite of robust.
Holocene: The epoch of the Cenozoic Era starting around 12,000 years ago and lasting arguably through the present.
Ice-Free Corridor model: Theory that the first Native Americans crossed to the Americas through a passage between glaciers.
Institutions: Long-lasting and influential cultural constructs. Examples include government, organized religion, academia, and the economy.
Last Glacial Maximum: The time 23,000 years ago when the most recent ice age was the most intense.
Later Stone Age: Time period following the Middle Stone Age with a diversification in tool types, starting around 50,000 years ago.
Levant: The eastern coast of the Mediterranean. The site of early modern human expansion from Africa and later one of the centers of agriculture.
Megafauna: Large ancient animals that may have been hunted to extinction by people around the world.
Mental eminence: The chin on the mandible of modern H. sapiens. One of the defining traits of our species.
Microlith: Small stone tool found in the Later Stone Age; also called a bladelet.
Middle Stone Age: Time period known for Mousterian lithics that connects African archaic to modern Homo sapiens.
Monumental architecture: Large and labor-intensive constructions that signify the power of the elite in a sedentary society. A common type is the pyramid, a raised crafted structure topped with a point or platform.
Mosaic: Composed from a mix or composite of traits.
Neolithic Revolution: Time of rapid change to human cultures due to the invention of agriculture, starting around 12,000 years ago.
Ochre: Iron-based mineral pigment that can be a variety of yellows, reds, and browns. Used by modern human cultures worldwide since at least 80,000 years ago.
Sahul: Ancient landmass connecting New Guinea and Australia.
Sedentarism: Lifestyle based on having a stable home area; the opposite of nomadism.
Southern Dispersal model: Theory that modern H. sapiens expanded from East Africa by crossing the Red Sea and following the coast east across Asia.
Subsistence strategy: The method an organism uses to find nourishment and other resources.
Sunda: Ancient Asian landmass that incorporated modern Southeast Asia.
Supraorbital torus: The bony brow ridge across the top of the eye orbits on many hominin crania.
Upper Paleolithic: Time period considered synonymous with the Later Stone Age.
Urbanization: The increase of population density as people settled together in cities.
Wallacea: Archipelago southeast of Sunda with different biodiversity than Asia.
Younger Dryas: The rapid change in global climate—notably a cooling of the Northern Hemisphere—13,000 years ago.
For Further Exploration
Websites
First-person virtual tour of Lascaux cave with annotated cave art: Ministère de la Culture and Musée d’Archéologie Nationale. “Visit the cave” Lascaux website.
Online anthropology magazine articles related to paleoanthropology and human evolution: SAPIENS. “Evolution.” SAPIENS website.
Various presentations of information about hominin evolution: Smithsonian Institution. “What does it mean to be human?” Smithsonian National Museum of Natural History website.
Magazine-style articles on archaeology and paleoanthropology: ThoughtCo. “Archaeology.” ThoughtCo. Website.
Database of comparisons across hominins and primates: University of California, San Diego. “MOCA Domains.” Center for Academic Research & Training in Anthropogeny website.
Books
Engaging book that covers human-made changes to the environment with industrialization and globalization: Kolbert, Elizabeth. 2014. The Sixth Extinction: An Unnatural History. New York: Bloomsbury.
Overview of what human life was like among the environmental shifts of the Ice Age: Woodward, Jamie. 2014. The Ice Age: A Very Short Introduction. Oxford: OUP Press.
Articles
Recent review paper about the current state of paleoanthropology research: Stringer, C. 2016. “The Origin and Evolution of Homo sapiens.” Philosophical Transactions of the Royal Society B 371 (1698).
Overview of the history of American paleoanthropology and the many debates that have occurred over the years: Trinkaus, E. 2018. “One Hundred Years of Paleoanthropology: An American Perspective.” American Journal of Physical Anthropology 165 (4): 638–651.
Amazing magazine article that synthesizes hominin evolution and why it is important to study this subject: Wheelwright, Jeff. 2015. “Days of Dysevolution.” Discover 36 (4): 33–39.
Fascinating research on Ötzi, a mummy from 5,000 years ago: Wierer, Ursula, Simona Arrighi, Stefano Bertola, Günther Kaufmann, Benno Baumgarten, Annaluisa Pedrotti, Patrizia Pernter, and Jacques Pelegrin. 2018. “The Iceman’s Lithic Toolkit: Raw Material, Technology, Typology and Use.” PLOS One 13 (6): e0198292. https://doi.org/10.1371/journal.pone.0198292.
Documentaries
PBS NOVA series covering the expansion of modern Homo sapiens and interbreeding with archaic humans: Brown, Nicholas, dir. 2015. First Peoples. Edmonton: Wall to Wall Television. Amazon Prime Video.
PBS NOVA special featuring the footprints found in White Sands National Park: Falk, Bella, dir. 2016. Ice Age Footprints. Boston: Windfall Films. https://www.pbs.org/wgbh/nova/video/ice-age-footprints/.
PBS NOVA special about how modern humans evolved adaptations to different environments. Shows how present-day people live around the world: Thompson, Niobe, dir. 2016. Great Human Odyssey. Edmonton: Clearwater Documentary. https://www.pbs.org/wgbh/nova/evolution/great-human-odyssey.html.
References
Araujo, Bernardo B. A., Luiz Gustavo R. Oliveira-Santos, Matheus S. Lima-Ribeiro, José Alexandre F. Diniz-Filho, and Fernando A. S. Fernandez. 2017. “Bigger Kill Than Chill: The Uneven Roles of Humans and Climate on Late Quaternary Megafaunal Extinctions.” Quaternary International 431: 216–222.
Armelagos, George J., Peter J. Brown, and Bethany Turner. 2005. “Evolutionary, Historical, and Political Economic Perspectives on Health and Disease.” Social Science & Medicine 61 (4): 755–765.
Armstrong, C. G., J. E. D. Miller, A. C. McAlvay, P. M. Ritchie, and D. Lepofsky. 2021. “Historical Indigenous Land-Use Explains Plant Functional Trait Diversity. Ecology and Society 26 (2): 6.
Bar-Yosef Mayer, Daniella E., Bernard Vandermeersch, and Ofer Bar-Yosef. 2009. “Shells and Ochre in Middle Paleolithic Qafzeh Cave, Israel: Indications for Modern Behavior.” Journal of Human Evolution 56 (3): 307–314.
Barbetti, M., and H. Allen. 1972. “Prehistoric Man at Lake Mungo, Australia, by 32,000 Years Bp.” Nature 240 (5375): 46–48.
Bennett, M. R., D. Bustos, J. S. Pigati, K. B. Springer, T. M. Urban, V. T. Holliday, Sally C. Reynolds, et al. (2021). “Evidence of Humans in North America during the Last Glacial Maximum.” Science 373 (6562): 1528–1531.
Bowler, J. M., Rhys Jones, Harry Allen, and A. G. Thorne. 1970. “Pleistocene Human Remains from Australia: A Living Site and Human Cremation from Lake Mungo, Western New South Wales.” World Archaeology 2 (1): 39–60.
Brown, Peter. 1999. “The First Modern East Asians? Another Look at Upper Cave 101, Liujiang and Minatogawa 1.” In Interdisciplinary Perspectives on the Origins of the Japanese, edited by K. Omoto, 105–131. Kyoto: International Research Center for Japanese Studies.
Brown, Peter. 2000. “Australian Pleistocene Variation and the Sex of Lake Mungo 3.” Journal of Human Evolution 38 (5): 743–749.
Clarkson, Chris, Zenobia Jacobs, Ben Marwick, Richard Fullagar, Lynley Wallis, Mike Smith, Richard G. Roberts, et al. 2017. “Human Occupation of Northern Australia by 65,000 Years Ago.” Nature 547 (7663): 306–310.
Cohen, Mark Nathan. 1977. The Food Crisis in Prehistory: Overpopulation and the Origins of Agriculture. New Haven, CT: Yale University Press.
Cohen, Mark Nathan, and George J. Armelagos, eds. 1984. Paleopathology at the Origins of Agriculture. Orlando, FL: Academic Press.
Cohen, Mark Nathan, and Gillian M. M. Crane-Kramer, eds. 2007. Ancient Health: Skeletal Indicators of Agricultural and Economic Intensification. Gainesville, FL: University Press of Florida.
Copes-Gerbitz, K., S. Hagerman, and L. Daniels. 2021. “Situating Indigenous Knowledge for Resilience in Fire-Dependent Social-Ecological Systems.” Ecology and Society 26(4): 25. https://www.ecologyandsociety.org/vol26/iss4/art25/.
Coqueugniot, Hélène, Olivier Dutour, Baruch Arensburg, Henri Duday, Bernard Vandermeersch, and Anne-Marie Tillier. 2014. “Earliest Cranio-Encephalic Trauma from the Levantine Middle Palaeolithic: 3-D Reappraisal of the Qafzeh 11 Skull, Consequences of Pediatric Brain Damage on Individual Life Condition and Social Care.” PLOS ONE 9 (7): e102822.
Crittenden, Alyssa N., and Stephanie L. Schnorr. 2017. “Current Views on Hunter‐Gatherer Nutrition and the Evolution of the Human Diet.” American Journal of Physical Anthropology 162 (S63): 84–109.
d’Errico, Francesco, Lucinda Backwell, Paola Villa, Ilaria Degano, Jeannette J. Lucejko, Marion K. Bamford, Thomas F. G. Higham, Maria Perla Colombini, and Peter B. Beaumont. 2012. “Early Evidence of San Material Culture Represented by Organic Artifacts from Border Cave, South Africa.” Proceedings of the National Academy of Sciences 109 (33): 13214–13219.
d’Errico, Francesco, Christopher Henshilwood, Marian Vanhaeren, and Karen Van Niekerk. 2005. “Nassarius Kraussianus Shell Beads from Blombos Cave: Evidence for Symbolic Behaviour in the Middle Stone Age.” Journal of Human Evolution 48 (1): 3–24.
Dannemann, Michael, and Fernando Racimo. 2018. “Something Old, Something Borrowed: Admixture and Adaptation in Human Evolution.” Current Opinion in Genetics & Development 53: 1–8.
Day, M. H. 1969. “Omo Human Skeletal Remains.” Nature 222: 1135–1138.
Dillehay, Tom D., Carlos Ocampo, José Saavedra, Andre Oliveira Sawakuchi, Rodrigo M. Vega, Mario Pino, Michael B. Collins, et al. 2015. “New Archaeological Evidence for an Early Human Presence at Monte Verde, Chile.” PLOS ONE 10 (11): e0141923. doi:10.1371/journal.pone.0141923.
Dow, Gregory K., Clyde G. Reed, and Nancy Olewiler. 2009. “Climate Reversals and the Transition to Agriculture.” Journal of Economic Growth 14 (1): 27–53.
Durband, Arthur C. 2014. “Brief Communication: Artificial Cranial Modification in Kow Swamp and Cohuna.” American Journal of Physical Anthropology 155 (1): 173–178.
Ember, Carol R. N.d. “Hunter-Gatherers.” Explaining Human Culture. Human Relations Area Files. Accessed March 4, 2023. https://hraf.yale.edu/ehc/summaries/hunter-gatherers.
Erlandson, Jon M., Todd J. Braje, Kristina M. Gill, and Michael H. Graham. 2015. “Ecology of the Kelp Highway: Did Marine Resources Facilitate Human Dispersal from Northeast Asia to the Americas?” The Journal of Island and Coastal Archaeology 10 (3): 392–411.
Fladmark, K. R. 1979. “Routes: Alternate Migration Corridors for Early Man in North America.” American Antiquity 44 (1): 55–69.
Fletcher, M. S., T. Hall, and A. N. Alexandra. 2021. “The Loss of an Indigenous Constructed Landscape Following British Invasion of Australia: An Insight into the Deep Human Imprint on the Australian Landscape.” Ambio 50(1): 138–149.
Fu, Qiaomei, Mateja Hajdinjak, Oana Teodora Moldovan, Silviu Constantin, Swapan Mallick, Pontus Skoglund, Nick Patterson, et al. 2015. “An Early Modern Human from Romania with a Recent Neanderthal Ancestor.” Nature 524 (7564): 216–219.
Fuller, Dorian Q. 2010. “An Emerging Paradigm Shift in the Origins of Agriculture.” General Anthropology 17 (2): 1, 8–11.
Gammage, B. 2008. “Plain Facts: Tasmania under Aboriginal Management.” Landscape Research 33 (2): 241–254.
Germonpré, Mietje, Martina Lázničková-Galetová, and Mikhail V. Sablin. 2012. “Palaeolithic Dog Skulls at the Gravettian Předmostí Site, the Czech Republic.” Journal of Archaeological Science 39 (1): 184–202.
Gröning, Flora, Jia Liu, Michael J. Fagan, and Paul O’Higgins. 2011. “Why Do Humans Have Chins? Testing the Mechanical Significance of Modern Human Symphyseal Morphology with Finite Element Analysis.” American Journal of Physical Anthropology 144 (4): 593–606.
Harvati, Katerina. 2009. “Into Eurasia: A Geometric Morphometric Reassessment of the Upper Cave (Zhoukoudian) Specimens.” Journal of Human Evolution 57 (6): 751–762.
Headland, Thomas N., Lawrence A. Reid, M. G. Bicchieri, Charles A. Bishop, Robert Blust, Nicholas E. Flanders, Peter M. Gardner, Karl L. Hutterer, Arkadiusz Marciniak, and Robert F. Schroeder. 1989. “Hunter-Gatherers and Their Neighbors from Prehistory to the Present.” Current Anthropology 30 (1): 43–66.
Henshilwood, Christopher S., Francesco d’Errico, Karen L. van Niekerk, Yvan Coquinot, Zenobia Jacobs, Stein-Erik Lauritzen, Michel Menu, and Renata García-Moreno. 2011. “A 100,000-Year-Old Ochre-Processing Workshop at Blombos Cave, South Africa.” Science 334 (6053): 219–222.
Hershkovitz, Israel, Gerhard W. Weber, Rolf Quam, Mathieu Duval, Rainer Grün, Leslie Kinsley, Avner Ayalon, et al. 2018. “The Earliest Modern Humans Outside Africa.” Science 359 (6374): 456–459.
Hublin, Jean-Jacques, Abdelouahed Ben-Ncer, Shara E. Bailey, Sarah E. Freidline, Simon Neubauer, Matthew M. Skinner, Inga Bergmann, et al. 2017. “New Fossils from Jebel Irhoud, Morocco, and the Pan-African Origin of Homo sapiens.” Nature 546 (7657): 289–292.
Lepofsky, D., N. F. Smith, N. Cardinal, J. Harper, M. Morris, M., Gitla (Elroy White), Randy Bouchard, et al. 2015. “Ancient Shellfish Mariculture on the Northwest Coast of North America.” American Antiquity 80 (2): 236–259.
Lieberman, Daniel E. 2015. “Human Locomotion and Heat Loss: An Evolutionary Perspective.” Comprehensive Physiology 5 (1): 99–117.
Lieberman, Daniel E., Brandeis M. McBratney, and Gail Krovitz. 2002. “The Evolution and Development of Cranial Form in Homo sapiens.” Proceedings of the National Academy of Sciences 99 (3): 1134–1139.
Lieberman, Daniel E., Osbjorn M. Pearson, and Kenneth M. Mowbray. 2000. “Basicranial Influence on Overall Cranial Shape.” Journal of Human Evolution 38 (2): 291–315.
Liu, Wu, María Martinón-Torres, Yan-jun Cai, Song Xing, Hao-wen Tong, Shu-wen Pei, Mark Jan Sier, Xiao-hong Wu, R. Lawrence Edwards, and Hai Cheng. 2015. “The Earliest Unequivocally Modern Humans in Southern China.” Nature 526 (7575): 696-699.
Lucas, Peter W. 2007. “The Evolution of the Hominin Diet from a Dental Functional Perspective.” In Evolution of the Human Diet: The Known, the Unknown, and the Unknowable, edited by Peter S. Ungar, 31–38 Oxford, UK: Oxford University Press.
Marsh, W. A. & Bello, S. (2023). Cannibalism and burial in the late Upper Palaeolithic: Combining archaeological and genetic evidence. Quaternary Science Reviews, 319, 108309. https://doi.org/10.1016/j.quascirev.2023.108309
McCarthy, Robert C., and Lynn Lucas. 2014. “A Morphometric Reassessment of Bou-Vp-16/1 from Herto, Ethiopia.” Journal of Human Evolution 74: 114–117.
McDougall, Ian, Francis H. Brown, and John G. Fleagle. 2005. “Stratigraphic Placement and Age of Modern Humans from Kibish, Ethiopia.” Nature 433 (7027): 733–736.
McNiven, I. J., J. Crouch, T. Richards, N. Dolby, and G. Jacobsen. 2012. “Dating Aboriginal Stone-Walled Fishtraps at Lake Condah, Southeast Australia.” Journal of Archaeological Science 39 (2): 268–286.
McNiven, I., J. Crouch, T. Richards, K. Sniderman, N. Dolby, and G. Mirring. 2015. “Phased Redevelopment of an Ancient Gunditjmara Fish Trap over the Past 800 Years: Muldoons Trap Complex, Lake Condah, Southwestern Victoria.” Australian Archaeology 81 (1): 44–58.
Michel, Véronique, Hélène Valladas, Guanjun Shen, Wei Wang, Jian-xin Zhao, Chuan-Chou Shen, Patricia Valensi, and Christopher J. Bae. 2016. “The Earliest Modern Homo sapiens in China?” Journal of Human Evolution 101: 101–104.
Miller, D. Shane, Vance T. Holliday, and Jordon Bright. 2013. “Clovis across the Continent.” In Paleoamerican Odyssey, edited by Kelly E. Graf, Caroline V. Ketron, and Michael R. Waters, 207–220. College Station: Texas A&M University Press.
Neubauer, Simon, Jean-Jacques Hublin, and Philipp Gunz. 2018. “The Evolution of Modern Human Brain Shape.” Science Advances 4 (1): eaao5961. https://doi.org/10.1126/sciadv.aao5961.
Pearson, Osbjorn M. 2000. “Postcranial Remains and the Origin of Modern Humans.” Evolutionary Anthropology 9: 229–247.
Pearson, Osbjorn M. 2008. “Statistical and Biological Definitions of ‘Anatomically Modern’ Humans: Suggestions for a Unified Approach to Modern Morphology.” Evolutionary Anthropology: Issues, News, and Reviews 17 (1): 38–48.
Pobiner, B., Pante, M. & Keevil, T. (2023). Early Pleistocene cut marked hominin fossil from Koobi Fora, Kenya. Sci Rep, 13, 9896. https://doi.org/10.1038/s41598-023-35702-7
Pietschnig, Jakob, Lars Penke, Jelte M. Wicherts, Michael Zeiler, and Martin Voracek. 2015. “Meta-Analysis of Associations between Human Brain Volume and Intelligence Differences: How Strong Are They and What Do They Mean?” Neuroscience & Biobehavioral Reviews 57: 411–432.
Posth, Cosimo, Nathan Nakatsuka, Iosif Lazaridis, Pontus Skoglund, Swapan Mallick, Thiseas C. Lamnidis, Nadin Rohland, et al. 2018. “Reconstructing the Deep Population History of Central and South America.” Cell 175 (5): 1185–1197.
Potter, Ben A., James F. Baichtal, Alwynne B. Beaudoin, Lars Fehren-Schmitz, C. Vance Haynes, Vance T. Holliday, Charles E. Holmes, et al. 2018. “Current Evidence Allows Multiple Models for the Peopling of the Americas.” Science Advances 4 (8): eaat5473. https://doi.org/10.1126/sciadv.aat5473.
Reich, David, Richard E. Green, Martin Kircher, Johannes Krause, Nick Patterson, Eric Y. Durand, Bence Viola, et al. 2010. “Genetic History of an Archaic Hominin Group from Denisova Cave in Siberia.” Nature 468 (7327): 1053–1060.
Reich, David, Nick Patterson, Martin Kircher, Frederick Delfin, Madhusudan R. Nandineni, Irina Pugach, Albert Min-Shan Ko, et al. 2011. “Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania.” American Journal of Human Genetics 89 (4): 516–528.
Richter, Daniel, Rainer Grün, Renaud Joannes-Boyau, Teresa E. Steele, Fethi Amani, Mathieu Rué, Paul Fernandes, et al. 2017. “The Age of the Hominin Fossils from Jebel Irhoud, Morocco, and the Origins of the Middle Stone Age.” Nature 546 (7657): 293–296.
Roberts, Patrick, and Brian A. Stewart. 2018. “Defining the ‘Generalist-Specialist’ Niche for Pleistocene Homo sapiens.” Nature Human Behaviour 2: 542–550.
Rougier, Helene, Ştefan Milota, Ricardo Rodrigo, Mircea Gherase, Laurenţiu Sarcinǎ, Oana Moldovan, João Zilhão, et al. 2007. “Peştera Cu Oase 2 and the Cranial Morphology of Early Modern Europeans.” Proceedings of the National Academy of Sciences 104 (4): 1165–1170.
Sahlins, Marshall. 1968. “Notes on the Original Affluent Society.” In Man the Hunter, edited by R. B. Lee and I. DeVore, 85–89. New York: Aldine Publishing Company.
Saladié, P. & Rodríguez-Hidalgo, A. (2017). Archaeological Evidence for Cannibalism in Prehistoric Western Europe: from Homo antecessor to the Bronze Age. Journal of Archaeological Method and Theory 24, 1034–1071. https://doi.org/10.1007/s10816-016-9306-y
Sawyer, G. J., and Blaine Maley. 2005. “Neanderthal Reconstructed.” The Anatomical Record (Part B: New Anat.) 283 (1): 23–31.
Scerri, Eleanor M. L., Mark G. Thomas, Andrea Manica, Philipp Gunz, Jay T. Stock, Chris Stringer, Matt Grove, et al. 2018. “Did Our Species Evolve in Subdivided Populations Across Africa, and Why Does It Matter?” Trends in Ecology & Evolution 33 (8): 582–594.
Shea, John J. 2011. “Refuting a Myth about Human Origins.” American Scientist 99 (2): 128–135.
Shea, John J., and Ofer Bar-Yosef. 2005. “Who Were the Skhul/Qafzeh People? An Archaeological Perspective on Eurasia’s Oldest Modern Humans.” Journal of the Israel Prehistoric Society 35: 451–468.
Slatkin, Montgomery, and Fernando Racimo. 2016. “Ancient DNA and Human History.” Proceedings of the National Academy of Sciences 113 (23): 6380–6387.
Smithsonian Institution. (2023, June 26). Humans’ evolutionary relatives butchered one another 1.45 million years ago. Smithsonian Institution. https://www.si.edu/newsdesk/releases/humans-evolutionary-relatives-butchered-one-another-145-million-years-ago
Smith, Fred H., James C. M. Ahern, Ivor Janković, and Ivor Karavanić. 2017. “The Assimilation Model of Modern Human Origins in Light of Current Genetic and Genomic Knowledge.” Quaternary International 450: 126–136.
Smith, Michael. 2009. “V. Gordon Childe and the Urban Revolution: A Historical Perspective on a Revolution in Urban Studies.” Town Planning Review 80 (1): 3–29.
Spence, M., & Jackson, L. J. (2014). The Bioarchaeology of Cannibalism at the Charity Site. Journal of The Ontario Archaeological Society , 94, 65–80. https://www.academia.edu/14371786/The_Bioarchaeology_of_Cannibalism_at_the_Charity_Site
Stock, Jay T. 2008. “Are Humans Still Evolving?” EMBO Reports 9 (Suppl 1): S51–S54.
Swisher, Mark E., Dennis L. Jenkins, Lionel E. Jackson Jr., and Fred M. Phillips. 2013. “A Reassessment of the Role of the Canadian Ice-Free Corridor in Light of New Geological Evidence.” Poster Symposium 5B: Geology, Geochronology and Paleoenvironments of the First Americans at the Paleoamerican Odyssey Conference, Santa Fe, New Mexico, October 16–19.
Thorne, A. G., and P. G. Macumber. 1972. “Discoveries of Late Pleistocene Man at Kow Swamp, Australia.” Nature 238 (5363): 316–319.
Trinkaus, Erik, Ştefan Milota, Ricardo Rodrigo, Gherase Mircea, and Oana Moldovan. 2003a. “Early Modern Human Cranial Remains from the Peştera Cu Oase, Romania.” Journal of Human Evolution 45 (3): 245–253.
Trinkaus, Erik, Oana Moldovan, Adrian Bîlgăr, Laurenţiu Sarcina, Sheela Athreya, Shara E Bailey, Ricardo Rodrigo, Gherase Mircea, Thomas Higham, and Christopher Bronk Ramsey. 2003b. “An Early Modern Human from the Peştera Cu Oase, Romania.” Proceedings of the National Academy of Sciences 100 (20): 11231–11236.
Ullrich, H. (2005). Cannibalistic rites within mortuary practices from the palaeolithic to middle aged in Europe. Anthropologie (1962-), 43(2/3), 249–261. http://www.jstor.org/stable/26292739
Velemínská, J., J. Brůzek, P. Velemínský, L. Bigoni, A. Sefcáková, and S. Katina. 2008. “Variability of the Upper-Palaeolithic Skulls from Predmostí Near Prerov (Czech Republic): Craniometric Comparison with Recent Human Standards.” Homo 59 (1): 1–26.
Vidal, Céline M., Christine S. Lane, Asfawossen Asrat, Dan N. Barfod, Darren F. Mark, Emma L. Tomlinson, Ambdemichael Zafu Tadesse, et al. (2022). “Age of the Oldest Known Homo sapiens from Eastern Africa. Nature 601 (7894): 579–583.
Villa, P., Bouville, C., Courtin, J., Helmer, D., Mahieu, E., Shipman, P., Belluomini, G. & Branca, M. (1986). Cannibalism in the Neolithic. Science, 233(4762), 431–437. doi:10.1126/science.233.4762.431
Villa, Paola, Sylvain Soriano, Tsenka Tsanova, Ilaria Degano, Thomas F. G. Higham, Francesco d’Errico, Lucinda Backwell, Jeannette J. Lucejko, Maria Perla Colombini, and Peter B. Beaumont. 2012. “Border Cave and the Beginning of the Later Stone Age in South Africa.” Proceedings of the National Academy of Sciences 109 (33): 13208–13213.
Wall, Jeffrey D., and Deborah Yoshihara Caldeira Brandt. 2016. “Archaic Admixture in Human History.” Current Opinion in Genetics & Development 41: 93–97.
White, Tim D., Berhane Asfaw, David DeGusta, Henry Gilbert, Gary D. Richards, Gen Suwa, and F. Clark Howell. 2003. “Pleistocene Homo sapiens from Middle Awash, Ethiopia.” Nature 423 (6941): 742–747.
Woo, Ju-Kang. 1959. “Human Fossils Found in Liukiang, Kwangsi, China.” Vertebrata PalAsiatica 3 (3): 109–118.
Wu, XiuJie, Wu Liu, Wei Dong, JieMin Que, and YanFang Wang. 2008. “The Brain Morphology of Homo Liujiang Cranium Fossil by Three-Dimensional Computed Tomography.” Chinese Science Bulletin 53 (16): 2513–2519.
Acknowledgments
I could not have undertaken this project without the help of many who got me to where I am today. I extend sincere thank yous to the many colleagues and former students who have inspired me to keep learning and talking about anthropology. Thank you also to all who are involved in this textbook project. The anonymous reviewers truly sparked improvements to the chapter. Lastly, the staff of Starbucks #5772 also contributed immensely to this text.
Jonathan Marks, Ph.D., University of North Carolina at Charlotte
Adam P. Johnson, M.A., University of North Carolina at Charlotte/University of Texas at San Antonio
Student contributors to this chapter: Daphnée-Tiffany Kirouac Millan, Davina Paradis, Jung Jin Kim, and Nathan Dennis
This chapter is an adaptation of "Chapter 2: Evolution” by Jonathan Marks. In Explorations: An Open Invitation to Biological Anthropology, first edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0.
Learning Objectives
- Explain the relationship among genes, bodies, and organismal change.
- Discuss the shortcomings of simplistic understandings of genetics.
- Describe what is meant by the "biopolitics of heredity."
- Discuss issues caused by misuse of ideas about adaptations and natural selection.
- Examine and correct myths about evolution.
The Human Genome Project, an international initiative launched in 1990, sought to identify the entire genetic makeup of our species. For many scientists, it meant trying to understand the genetic underpinnings of what made humans uniquely human. James Watson, a codiscoverer of the helical shape of DNA, wrote that “when finally interpreted, the genetic messages encoded within our DNA molecules will provide the ultimate answers to the chemical underpinnings of human existence” (Watson 1990, 248). The underlying message is that what makes humans unique can be found in our genes. The Human Genome Project hoped to find the core of who we are and where we come from.
Despite its lofty goal, the Human Genome Project—even after publishing the entire human genome in January 2022—could not fully account for the many factors that contribute to what it is to be human. Richard Lewontin, Steven Rose, and Leon Kamin (2017) argue that genetic determinism of the sort assumed by the Human Genome Project neglects other essential dimensions that contribute to the development and evolution of human bodies, not to mention the role that culture plays. They use an apt metaphor of a cake to illustrate the incompleteness of reductive models. Consider the flavor of a cake and think of the ingredients listed in the recipe. The recipe includes ingredients such as flour, sugar, shortening, vanilla extract, eggs, and milk. Does raw flour taste like cake? Does sugar, vanilla extract, or any of the other ingredients taste like cake? They do not, and knowing the individual flavors of each ingredient does not tell us much about what cake tastes like. Even mixing all of the ingredients in the correct proportions does not get us cake. Instead, external factors such as baking at the right temperature, for the right amount of time, and even the particularities of our evolved sense of taste and smell are all necessary components of experiencing the cake. Lewontin, Rose, and Kamin (2017) argue that the same is true for humans and other organisms.
Knowing everything about cake ingredients does not allow us to fully know cake. Equally so, knowing everything about the genes found in our DNA does not allow us to fully know humans. Different, interacting levels are implicated in the development and evolution of all organisms, including humans. Genes, the structure of chromosomes, developmental processes, epigenetic tags, environmental factors, and still-other components all play key roles such that genetically reductive models of human development and evolution are woefully inadequate.
The complex interactions across many levels—genetic, developmental, and environmental—explain why we still do not know how our one-dimensional DNA nucleotide sequence results in a four-dimensional organism. This was the unfulfilled promise of the inception of the Human Genome Project in the 1980s and 1990s: the project produced the complete DNA sequence of a human cell in the hopes that it would reveal how human bodies are built and how to cure them when they are built poorly. Yet, that information has remained elusive. Presumably, the knowledge of how organisms are produced from DNA sequences will one day permit us to reconcile the discrepancies between patterns in anatomical evolution and molecular evolution.
In this chapter, we will consider multilevel evolution and explore evolution as a complex interaction between genetic and epigenetic factors as well as the environments in which organisms live. Next, we will examine the biopolitical nature of human evolution. We will then investigate problems that arise from attributing all traits to an adaptive function. Finally, we will address common misconceptions about evolution. The goal of this chapter is to provide you with the necessary toolkit for understanding the molecular, anatomical, and political dimensions of evolution.
Evolution Happens at Multiple Levels
Following Richard Dawkins’s publication of The Selfish Gene in 1976, the scientific imagination was captured by the potential of genomics to reveal how genes are copied by Darwinian selection. Dawkins argues that the genes in individuals that contribute to greater reproductive success are the units of selection. His conception of evolution at the molecular level undercuts the complex interactions between organisms and their environments, which are not expressed genomically but are nevertheless key drivers in evolution.
By the 1980s, the acknowledgment among most biologists that even though genes construct bodies, genes and bodies evolve at different rates and with distinct patterns. This realization led to a renewed focus on how bodies change. The Evolutionary Synthesis of the 1930s–1970s had reduced organisms to their genotypes and species to their gene pools, which provided valuable insights about the processes of biological change, but it was only a first approximation. Animals are in fact reactive and adaptable beings, not passive and inert genotypes. Species are clusters of socially interacting and reproductively compatible organisms.

Once we accept that evolutionary change is fundamentally genetic change, we can ask: How do bodies function and evolve? How do groups of animals come to see one another as potential mates or competitors for mates, as opposed to just other creatures in the environment? Are there evolutionary processes that are not explicable by population genetics? These questions—which lead us beyond reductive assumptions—were raised in the 1980s by Stephen Jay Gould, the leading evolutionary biologist of the late 20th century (see: Gould 2003; 1996).
Gould spearheaded a movement to identify and examine higher-order processes and features of evolution that were not adequately explained by population genetics. For example, extinction, which was such a problem for biologists of the 1600s, could now be seen as playing a more complex role in the history of life than population genetics had been able to model. Gould recognized that there are two kinds of extinctions, each with different consequences: background extinctions and mass extinctions. Background extinctions are those that reflect the balance of nature, because in a competitive Darwinian world, some things go extinct and other things take their place. Ecologically, your species may be adapted to its niche, but if another species comes along that’s better adapted to the same niche, eventually your species will go extinct. It sucks, but it is the way of all life: you come into existence, you endure, and you pass out of existence. But mass extinctions are quite different. They reflect not so much the balance of nature as the wholesale disruption of nature: many species from many different lineages dying off at roughly the same time—presumably as the result of some kind of rare ecological disaster. The situation may not be survival of the fittest as much as survival of the luckiest. The result, then, would be an ecological scramble among the survivors. Having made it through the worst, the survivors could now simply divide up the new ecosystem amongst themselves, since their competitors were gone. Something like this may well have happened about 65 million years ago, when a huge asteroid hit the Yucatan Peninsula, which mammals survived but dinosaurs did not (Figure 3.1). Something like this may be happening now, due to human expansion and environmental degradation. Note, though, that there is only a limited descriptive role here for population genetics: the phenomena we are describing are about organisms and species in ecosystems.
Another question involved the disconnect between properties of species and the properties of gene pools. For example, there are upwards of 15 species of gibbons but only two species of chimpanzees. Why? There are upwards of 20 species of guenons but fewer than ten of baboons. Why? Are there genes for that? It seems unlikely. Gould suggested that species, as units of nature, might have properties that are not reducible to the genes in their cells. For example, rates of speciation and extinction might be properties of their ecologies and histories rather than their genes. Thus, relationships between environmental contexts and variability within a species result in degrees of resistance to extinction and affect the frequency and rates at which clades diversify (Lloyd & Gould 1993). Consistent biases of speciation rates might well produce patterns of macroevolutionary diversity that are difficult to explain genetically and better understood ecologically. Gould called such biases in speciation rates species selection—a higher-order process that invokes competition between species, in addition to the classic Darwinian competition between individuals.
One of Gould’s most important studies involved the very nature of species. In the classical view, a species is continually adapting to its environment until it changes so much that it is a different species than it was at the beginning of this sentence (Eldredge & Gould 1972). That implies that the species is a fundamentally unstable entity through time, continuously changing to fit in. But suppose, argued Gould along with paleontologist Niles Eldredge, a species is more stable through time and only really adapts during periods of ecological instability and change. Then we might expect to find in the fossil record long equilibrium periods—a few million years or so—in which species don’t seem to change much, punctuated by relatively brief periods in which they change a bit and then stabilize again as new species. They called this idea punctuated equilibria. The idea helps to explain certain features of the fossil record, notably the existence of small anatomical “gaps” between closely related fossil forms (Figure 3.2). Its significance lies in the fact that although it incorporates genetics, punctuated equilibria is not really a theory of genetics but one of types bodies in deep time.
Punctuated equilibria is seen across taxa, with long periods in the fossil record representing little phenotypic change. These periods of stability are disrupted by shorter periods of rapid adaptation, the process through which populations of organisms become suited to living in their environments. Phenotypic changes are often coupled with drastic climatic or ecological changes that affect the milieu in which organisms live. For example, throughout much of hominin evolutionary history, brain size was closely associated with body size and thus remained mostly stable. However, changes occurred in average hominin brain size at around 100 thousand years ago, 1 million years ago, and 1.8 million years ago. Several hypotheses have been put forth to explain these changes, including unpredictability in climate and environment (Potts 1998), social development (Barton 1996), and the evolution of language (Deacon 1998). Evidence from the fossil record, paleoclimate models, and comparative anatomy suggests that the changes observed in hominin lineage result from biocultural processes—that is, the coalescence of environmental and cultural factors that selected for larger brains (Marks 2015; Shultz, Nelson, & Dunbar 2012).
In response to the call for a theory of the evolution of form, the field of evo-devo—the intersection of evolutionary and developmental biology—arose. The central focus here is on how changes in form and shape arise. An embryo matures by the stimulation of certain cells to divide, forming growth fields. The interactions and relationships among these growth fields generate the structures of the body. The hox genes that regulate these growth fields turn out to be highly conserved across the animal kingdom. This is because they repeatedly turn on and off the most basic genes guiding the animal’s development, and thus any changes to them would be catastrophic. Indeed, these genes were first identified by manipulating them in fruit flies, such that one could produce a bizarre mutant fruit fly that grew a pair of legs where its antennae were supposed to be (Kaufman, Seeger, and Olsen 1990).
Certain genetic changes can alter the fates of cells and the body parts, while other genetic changes can simply affect the rates at which neighboring groups of cells grow and divide, thus producing physical bumps or dents in the developing body. The result of altering the relationships among these fields of cellular proliferation in the growing embryo is allometry, or the differential growth of body parts. As an animal gets larger—either over the course of its life or over the course of macroevolution—it often has to change shape in order to live at a different size. Many important physiological functions depend on properties of geometric area: the strength of a bone, for example, is proportional to its cross-sectional area. But area is a two-dimensional quality, while growing takes place in three dimensions—as an increase in mass or volume. As an animal expands, its bones necessarily weaken, because volume expands faster than area does. Consequently a bigger animal has more stress on its bones than a smaller animal does and must evolve bones even thicker than they would be by simply scaling the animal up proportionally. In other words, if you expand a mouse to the size of an elephant, it will nevertheless still have much thinner bones than the elephant does. But those giant mouse bones will unfortunately not be adequate to the task. Thus, a giant mouse would have to change aspects of its form to maintain function at a larger size (see Figure 3.3).
Physiologically, we would like to know how the body “knows” when to turn on and off the genes that regulate growth to produce a normal animal. Evolutionarily, we would like to know how the body “learns” to alter the genetic on/off switch (or the genetic “slow down/speed up” switch) to produce an animal that looks different. Moreover, since organisms differ from one another, we would like to know how the developing body distinguishes a range of normal variation from abnormal variation. And, finally, how does abnormal variation eventually become normal in a descendant species?
Taking up these questions, Gould invoked the work of a British geneticist named Conrad H. Waddington, who thought about genetics in less reductive ways than his colleagues. Rather than isolate specific DNA sites to analyze their function, Waddington instead studied the inheritance of an organism’s reactivity—its ability to adapt to the circumstances of its life. In a famous experiment, he grew fruit fly eggs in an atmosphere containing ether. Most died, but a few survived somehow by developing a weird physical feature: a second thorax with a second pair of wings. Waddington bred these flies and soon developed a stable line of flies who would reliably develop a second thorax when grown in ether. Then he began to lower the concentration of ether, while continuing to selectively breed the flies that developed the strange appearance. Eventually he had a line of flies that would stably develop the “bithorax” phenotype–the suite of traits of an organism–even when there was no ether; it had become the “new normal.” The flies had genetically assimilated the bithorax condition.
Waddington was thus able to mimic the inheritance of acquired characteristics: what had been a trait stimulated by ether a few generations ago was now a normal part of the development of the descendants. Waddington recognized that while he had performed a selection experiment on genetic variants, he had not selected for particular traits. Rather, he helped produce the physiological tendency to develop particular traits when appropriately stimulated. He called that tendency plasticity and its converse, the tendency to stay the same even under weird environmental circumstances, canalization. Waddington had initially selected for plasticity, the tendency to develop the bithorax phenotype under weird conditions, and then, later, for canalization, the developmental normalization of that weird physical trait. Although Waddington had high stature in the community of geneticists, evolutionary biologists of the 1950s and 1960s regarded him with suspicion because he was not working within the standard mindset of reductionism, which saw evolution as the spread of genetic variants that coded for favorable traits. Both Waddington and Gould resisted contemporary intellectual paradigms that favored reductive accounts of evolutionary processes. They conceived of evolution as an emergent process in which many external factors (e.g. climate, environment, predation) and internal factors (e.g., genotypes, plasticity, canalization) coalesce to produce the evolutionary trends that we observe in the fossil record and our genome.
While Gould and Waddington both looked beyond the genome to understand evolution, the Human Genome Project—an international project with the goal of identifying each base pair in the human genome in the 1990s—generated a great deal of public interest in analyzing the human DNA sequence from the standpoint of medical genetics. Some of the rhetoric aimed to sell the public on investing a lot of money and resources in sequencing the human genome in order to show the genetic basis of heritable traits, cure genetic diseases, and learn what it means ultimately to be biologically human. However, the Human Genome Project was not actually able to answer those questions through the use of genetics alone, and thus a broader, more holistic account was required.
This holistic account came from decades of research in human biology and anthropology, which understood the human body as highly adaptable, dynamic, and emergent. For example, in the early 20th century, anthropologist Franz Boas measured the skulls of immigrants to the U.S., revealing that environmental, not merely genetic, factors affected skull shape. The growing human body adjusts itself to the conditions of life, such as diet, sunshine, high altitude, hard labor, population density, how babies are carried—any and all of which can have subtle but consistent effects upon its development. There can thus be no normal human form, only a context-specific range of human forms.
However, what the human biologists called human adaptability, evolutionary biologists called developmental plasticity, and evidence quickly began to mount for its cause being epigenetic modifications to DNA. Epigenetic modifications are changes to how genes are used by the body (as opposed to changes in the DNA sequences; see Chapter 4). Scientific interest shifted from the focus of the Human Genome Project to the ways that bodies are made by evolutionary-developmental processes, including epigenetics. What is meant by “epigenetic modification”? Evolution is about how descendants diverge from their ancestors. Inheritance from parent to offspring is still critical to this process, which occurs through genetic recombination: the pairing of homologous chromosomes and sharing of genetic material during meiosis (see Chapter 4). However, in the 21st century, the link between evolution and inheritance has broadened with a clearer understanding of how environmental and developmental factors shape bodies and the expression of genes, including epigenetic inheritance patterns. While offspring inherit their genes through random assortment during meiosis, environmental factors also shape how genes are used. When these epigenetic modifications occur in germ cells, they can be passed onto offspring. In these cases, there is no change in the DNA sequence but rather in how genes are used by the body due to DNA methylation and the structure of chromosomes due to histone acetylation (see Chapter 4).
In addition, we now recognize that evolution is affected by two other forms of intergenerational transmission and inheritance (in addition to genetics and epigenetics). These forms include behavioral variation and culture. That is, behavioral information can be transmitted horizontally (intragenerationally), permitting more rapid ways for organisms to adjust to the environment. And, then there is the fourth mode of transmission: the cultural or symbolic mode. It is proposed that humans are the only species that horizontally transmits an arbitrary set of rules to govern communication, social interaction, and thought. This shared information is symbolic and has resulted in what we recognize as “culture”: locally emergent worlds of names, words, pictures, classifications, revered pasts, possible futures, spirits, dead ancestors, unborn descendants, in-laws, politeness, taboo, justice, beauty, and story, all accompanied by practices and a material world of tools.
Consequently our contemporary ideas about evolution see the evolutionary processes as hierarchically organized and not restricted to the differential transmission of DNA sequences into the next generation. While that is indeed a significant part of evolution, the organism and species are nevertheless crucial to understanding how those DNA sequences get transmitted. Further, the transmission of epigenetic, behavioural, and symbolic information play a complex role in perpetuating our genes, bodies, and species. In the case of human evolution, one can readily see that symbolic information and cultural adaptation are far more central to our lives and our survival today than DNA and genetic adaptation. It is thus misleading to think of humans passively occupying an environmental niche. Rather, humans are actively engaged in constructing our own niches, as well as adapting to them and using them to adapt. The complex interplay between a species and its active engagement in creating its own ecology is known as niche construction. If we understand natural selection–the process by which populations adapt to their specific environments–as the effects that environmental context has on the reproductive success of organisms, then niche construction is the process through which organisms shape their own selective pressures.
Dig Deeper: Moving Beyond Genetic Determinism
Contemporary evolutionary biology and anthropology increasingly emphasize that genes operate within dynamic regulatory networks rather than acting as isolated determinants. As Carroll (2005) and Wray (2007) demonstrate, evolutionary change often arises not from mutations in structural genes but in their regulation—the timing, intensity, and location of gene expression. Such regulatory evolution can explain major anatomical and physiological innovations without invoking large genetic divergences. This view reframes evolution as an outcome of organizational complexity where genetic, developmental, and environmental processes intersect. This systems-level understanding also resonates with anthropological frameworks of biocultural embodiment, which recognize that social and ecological experiences can become biologically inscribed in the body. Meaney’s (2001) foundational epigenetic research focuses on maternal care in rats, presenting how nurturing behaviour modifies the expression of stress-response genes. This biological effect can persist into subsequent generations.
Recent human studies continue to expand this insight. Goldman & Sterner (2023) demonstrate how environmental exposures, inequality, and psychological stress influence the pace of biological aging, showing epigenetic modifications reflect the lived conditions of bodies over time. In Canada, this relationship between environment, history, and biology has profound implications. A 2023 scoping review on Canadian Indigenous populations and the epigenetic effects of intergenerational trauma (Schafte & Bruna, 2023) documents measurable biological patterns associated with colonial violence, displacement, and systemic inequity. By dissecting the obesity patterns in the Indigenous youth populations, the researchers present a clear connection between the parents who attended residential schools and biological health issues in their children years later. This holistic understanding of epigenetics shows an “embodied transmission of trauma and ill health across generations” (2023, p.9), underscoring that the effects of colonialism are not merely social but are biologically embodied, carried forward through mechanisms of gene regulation and stress physiology.
Understanding heredity as a process of interaction and regulation rather than genetic determinism opens the door to rethinking evolution as a flexible, context-driven phenomenon. Just as social experiences and ecological conditions can shape patterns of gene expression, environmental pressures can also influence the structure and behaviour of genomes across generations. This broader view of evolutionary change highlights the importance of considering mechanisms that fall outside of traditional, gradualist models.
The Biopolitics of Heredity
“Science isn’t political” is a sentiment that you have likely heard before. Science is supposed to be about facts and objectivity. It exists, or at least ought to, outside of petty human concerns. However, the sorts of questions we ask as scientists, the problems we choose to study, the categories and concepts we use, who gets to do science, and whose work gets cited are all shaped by culture. Doing science is a political act. This fact is markedly true for human evolution. While it is easier to create intellectual distance between us and fruit flies and viruses, there is no distance when we are studying ourselves. The hardest lesson to learn about human evolution is that it is intensely political. Indeed, to see it from the opposite side, as it were, the history of creationism—the belief that the universe was divinely created around 6,000 years ago—is essentially a history of legal decisions. For instance, in Tennessee v. John T. Scopes (1925), a schoolteacher was prosecuted for violating a law in Tennessee that prohibited the teaching of human evolution in public schools, where teachers were required by law to teach creationism.
More recently, legal decisions aimed at legislating science education have shaped how students are exposed to evolutionary theory. For instance, McLean v. Arkansas (1982) dispatched “scientific creationism” by arguing that the imposition of balanced teaching of evolution and creationism in science classes violates the Establishment Clause, separating church and state. Additionally, Kitzmiller v. Dover (Pennsylvania) Area School District (2005) dispatched the teaching of “intelligent design” in public school classrooms as it was deemed to not be science. In some cases, people see unbiblical things in evolution, although most Christian theologians are easily able to reconcile science to the Bible. In other cases, people see immoral things in evolution, although there is morality and immorality everywhere. And some people see evolution as an aspect of alt-religion, usurping the authority of science in schools to teach the rejection of the Christian faith, which would be unconstitutional due to the protected separation of church and state.
Clearly, the position that politics has nothing to do with science is untenable. But is the politics in evolution an aberration or is it somehow embedded in science? In the early 20th century, scientists commonly promoted the view that science and politics were separate: science was seen as a pure activity, only rarely corrupted by politics. And yet as early as World War I, the politics of nationalism made a hero of the German chemist Fritz Haber for inventing poison gas. And during World War II, both German doctors and American physicists, recruited to the war effort, helped to end many civilian lives. Therefore, we can think of the apolitical scientist as a self-serving myth that functions to absolve scientists of responsibility for their politics. The history of science shows how every generation of scientists has used evolutionary theory to rationalize political and moral positions. In the very first generation of evolutionary science, Darwin’s Origin of Species (1859) is today far more readable than his Descent of Man (1871). The reason is that Darwin consciously purged The Origin of Species of any discussion of people. And when he finally got around to talking about people, in The Descent of Man, he simply imbued them with the quaint Victorian prejudices of his age, and the result makes you cringe every few pages. There is plenty of politics in there—sexism, racism, and colonialism—because you cannot talk about people apolitically.
One immediate faddish deduction from Darwinism, popularized by Herbert Spencer (1864) as “survival of the fittest,” held that unfettered competition led to advancement in nature and to human history. Since the poor were purported losers in that struggle, anything that made their lives easier would go against the natural order. This position later came to be known ironically as “Social Darwinism.” Spencer was challenged by fellow Darwinian Thomas Huxley (1863), who agreed that struggle was the law of the jungle but observed that we don’t live in jungles anymore. The obligation to make lives better for others is a moral, not a natural, fact. We simultaneously inhabit a natural universe of descent from apes and a moral universe of injustice and inequality, and science is not well served by ignoring the latter.
Concurrently, the German biologist Ernst Haeckel’s 1868 popularization of Darwinism was translated into English a few years later as The History of Creation. As we saw earlier, Haeckel was determined to convince his readers that they were descended from apes, even in the absence of fossil evidence attesting to it. When he made non-Europeans into the missing links that connected his readers to the apes, and depicted them as ugly caricatures, he knew precisely what he was doing. Indeed, even when the degrading racial drawings were deleted from the English translation of his book, the text nevertheless made his arguments quite clear. And a generation later, when the Americans had not yet entered the Great War in 1916, a biologist named Vernon Kellogg visited the German High Command as a neutral observer and found that the officers knew a lot about evolutionary biology, which they had gotten from Haeckel and which rationalized their military aggressions. Kellogg went home and wrote a bestseller about it, called Headquarters Nights (1917). World War I would have been fought with or without evolutionary theory, but as a source of scientific authority, evolution—even if a perversion of the Darwinian theory—had very quickly attained global geopolitical relevance.
Oftentimes, politics in evolutionary science is subtle, due to the pervasive belief in the advancement of science. We recognize the biases of our academic ancestors and modify our scientific stories accordingly. But we can never be free of our own cultural biases, which are invisible to us, as much as our predecessors’ biases were invisible to them. In some cases, the most important cultural issues resurface in different guises each generation, like scientific racism. Scientific racism is the recruitment of science for the evil political ends of racism, and it has proved remarkably impervious to evolution. Before Darwin, there was creationist scientific racism, and after Darwin, there was evolutionist scientific racism. And there is still scientific racism today, self-justified by recourse to evolution, which means that scientists have to be politically astute and sensitive to the uses of their work to counter these social tendencies.
Consider this: Are you just your ancestry, or can you transcend it? If that sounds like a weird question, it was actually quite important to a turn-of-the-20th-century European society in which an old hereditary aristocracy was under increasing threat from a rising middle class. And that is why the very first English textbook of Mendelian genetics concluded with the thought that “permanent progress is a question of breeding rather than of pedagogics; a matter of gametes, not of training … the creature is not made but born” (Punnett 1905, 60). Translation: Not only do we now know a bit about how heredity works, but it’s also the most important thing about you. Trust me, I’m a scientist.
Yet evolution is about how descendants come to differ from ancestors. Do we really know that your heredity, your DNA, your ancestry, is the most important thing about you? That you were born, not made? After all, we do know that you could be born into slavery or as a peasant, and come from a long line of enslaved people or peasants, and yet not have slavery or peasantry be the most important thing about you. Whatever your ancestors were may unfortunately constrain what you can become, but as a moral precept, it should not. But just as science is not purely “facts and objectivity,” ancestry is not a strictly biological concept. Human ancestry is biopolitics, not biology.
Evolution is fundamentally a theory about ancestry, and yet ancestors are, in the broad anthropological sense, sacred: ancestors are often more meaningful symbolically than biologically. Just a few years after The Origin of Species (Darwin 1859), the British politician and writer Benjamin Disraeli declared himself to be on the side of the angels, not the apes, and to “repudiate with indignation and abhorrence those new-fangled theories” (Monypenny, Flavelle, and Buckle 1920, 105). He turned his back on an ape ancestry and looked to the angel; yet, he did so as a prominent Jew-turned-Anglican, who had personally transcended his humble roots and risen to the pinnacle of the Empire. Ancestry was certainly important, and Disraeli was famously proud of his, but it was also certainly not the most important thing, not the primary determinant of his place in the world. Indeed, quite the opposite: Disraeli’s life was built on the transcendence of many centuries of Jewish poverty and oppression in Europe. Humble ancestry was there to be superseded and nobility was there to be earned; Disraeli would later become the Earl of Beaconsfield. Clearly, “are you just your ancestry” is not a value-neutral question, and “the creature is not made, but born” is not a value-neutral answer.
Ancestry being the most important thing about a person became a popular idea twice in 20th century science. First, at the beginning of the century, when the eugenics movement in America called attention to “feeble-minded stocks,” which usually referred to the poor or to immigrants (see Figure 3.4). This movement culminated in Congress restricting the immigration of “feeble-minded races” (said to include Jews and Italians) in 1924, and the Supreme Court declaring it acceptable for states to sterilize their “feeble-minded” citizens involuntarily in 1927. After the Nazis picked up and embellished these ideas during World War II, Americans moved swiftly away from them in some contexts (e.g., for most people of European descent) while still strictly adhering in other contexts such as Japanese internment camps and immigration restrictions.

Ancestry again became paramount in the drumming up of public support for the Human Genome Project in the 1990s. Public support for sequencing the human genome was encouraged by a popular science campaign that featured books titled The Book of Man (Bodmer & McKie 1997), The Human Blueprint (Shapiro 1991), and The Code of Codes (Kevles & Hood 1993). These books generally promised cures for genetic diseases and a deeper understanding of the human condition. We can certainly identify progress in molecular genetics over the last couple of decades since the human genome was sequenced, but that progress has notably not been accompanied by cures for genetic diseases, nor by deeper understandings of the human condition.
Even at the most detailed and refined levels of genetic analysis, we still don’t have much of an understanding of the actual basis by which things seem to “run in families.” While the genetic basis of simple, if tragic, genetic diseases have become well-known—such as sickle-cell anemia, cystic fibrosis, and Tay-Sachs’ Disease—we still haven’t found the ostensible genetic basis for traits that are thought to have a strong genetic component. For example, a recent genetic summary found over 12,000 genetic sites that contributed to height yet still explained only about 40-50 percent of the variation in height among European ancestry but no more than 10-20 percent of variation of other ancestries, which we know strongly runs in families (Yengo Et al. 2022).
Partly in reaction to the reductionistic hype of the Human Genome Project, the study of epigenetics has become the subject of great interest. One famous natural experiment involves a Nazi-imposed famine in Holland over the winter of 1944–1945. Children born during and shortly after the famine experienced a higher incidence of certain health problems as adults, many decades later. Apparently, certain genes had been down-regulated early in development and remained that way throughout the course of life. Indeed, this modified regulation of the genes in response to the severe environmental conditions may have been passed on to their children.
Obviously one’s particular genetic constitution may play an important role in one’s life trajectory. But overvaluing that role may have important social and political consequences. In the first place, genotypes are rendered meaningful in a cultural universe. Thus, if you live in a strongly patriarchal society and are born without a Y chromosome (since human males are chromosomally XY and females XX), your genotype will indeed have a strong effect upon your life course. So even though the variation is natural, the consequences are political. The mediating factors are the cultural ideas about how people of different sexes ought to be treated, and the role of the state in permitting certain people to develop and thrive. More broadly, there are implications for public education if variation in intelligence is genetic. There are implications for the legal system if criminality is genetic. There are implications for the justice system if sexual preference, or sexual identity, is genetic. There are implications for the development of sports talent if that is genetic. And yet, even for the human traits that are more straightforward to measure and known to be strongly heritable, the DNA base sequence variation seems to explain little.
Genetic determinism or hereditarianism is the idea that “the creature is made, not born”—or, in a more recent formulation by James Watson, that “our fate is in our genes.” One of the major implications drawn from genetic determinism is that the feature in question must inevitably express itself; therefore, we can’t do anything about it. Therefore, we might as well not fund the social programs designed to ameliorate economic inequality and improve people’s lives, because their courses are fated genetically. And therefore, they don’t deserve better lives.
All of the “therefores” in the preceding paragraph are open to debate. What is important is that the argument relies on a very narrow understanding of the role of genetics in human life, and it misdirects the causes of inequality from cultural to natural processes. By contrast, instead of focusing on genes and imagining them to place an invisible limit upon social progress, we can study the ways in which your DNA sequence does not limit your capability for self-improvement or fix your place in a social hierarchy. In general, two such avenues exist. First, we can examine the ways in which the human body responds and reacts to environmental variation: human adaptability and plasticity. This line of research began with the anthropometric studies of immigrants by Franz Boas in the early 20th century and has now expanded to incorporate the epigenetic inheritance of modified human DNA. And second, we can consider how human lives are shaped by social histories—especially the structural inequalities within the societies in which they grow up.
Although it arises and is refuted every generation, the radical hereditarian position (genetic determinism) perennially claims to speak for both science and evolution. It does not. It is the voice of a radical fringe—perhaps naive, perhaps evil. It is not the authentic voice of science or of evolution. Indeed, keeping Charles Darwin’s name unsullied by protecting it from association with bad science often seems like a full-time job. Culture and epigenetics are very much a part of the human condition, and their roles are significant parts of the complete story of human evolution.
Special Topic: Oversexing the Gendered Body
While rapid mitochondrial evolution underscores the biological flexibility of organisms in response to environmental pressures, evolutionary theory is also shaped by another set of forces: cultural assumptions and social norms. Nowhere is this more visible than in scientific interpretations of sex and gender. Many modern gender roles stem from assumptions about sex differences that have accumulated throughout human history. While these roles may appear to be fixed stereotypes or biologically predetermined, they can be deconstructed by examining the processes of sexual selection through queer and feminist theoretical frameworks. Applying these lenses to evolutionary concepts allows for a deeper understanding of how cultural ideologies, particularly those surrounding gender and sexuality, shape interpretations of biological processes.
Darwin first introduced the concept of sexual selection in The Descent of Man (1871) to explain how males and females may have developed different traits that would be detrimental to the species’ overall survival (Vicedo, 2025). Unlike natural selection which is “selection by death,” sexual selection represents death by selection (Gayon, 2010). Darwin argued that males typically compete intrasexually for female attention, and that females exercise choice based on attractiveness or vigor, proving their fitness. However, when reframed through feminist theory, the amount of agency Darwin ascribed to females doesn’t reflect the societal assumptions surrounding gender roles in his era. Charlotte Perkins Gilman in her publication Women and Economics, argued that by the 1960s, men increasingly relied on social dominance over women rather than competition with other men (Vicedo, 2025). This dynamic required women to continually enhance their sexual appeal in exchange for economic security, a system she coined the “sexuo-economic relationship” (2025, p.5). This framework reveals the societal power imbalance between men and women, and how women are the ones sexualizing themselves and competing for partners, not men. Such processes would lead to the modern oversexualization of women.
Oversexualization, a cultural ideology that prioritizes sexual appeal over autonomy and well-being, further complicates interpretations of sexual selection. Brassard and company (2018) define oversexualization through four components: valuing people solely for their sexual appeal, societal norms of equating attractiveness with sexiness, sexual objectification, and the inappropriate imposition of sexuality (Brassard Et al., 2018, p.16-17). When oversexualization is observed within a population, it may signal that the pressures of sexual selection have intensified relative to that of natural selection, creating “excessive sex difference" (Vicedo, 2025). While many aspects of Gilman's arguments do not directly apply to contemporary gender dynamics, stereotypes rooted in historical gender expectations continue to shape women's experiences in the workforce and broader society (2025). Understanding sexual selection as a culturally mediated process, rather than as a simple competition amongst males, offers a more nuanced picture of how gender ideologies influence biological narratives. This intersection of culture and biology is crucial for studying gender roles, queer relationships, and sexual diversity across societies and time periods.
Adaptationism and the Panglossian Paradigm
The story of human evolution, and the evolution of all life for that matter, is never settled because evolution is ongoing. Additionally, because the conditions that shape evolutionary trajectories are not predetermined, evolution itself is emergent. Even during periods of ecological stability, when fewer macroevolutionary changes occur, populations of organisms continue to experience change. When ecological stability is disrupted, populations must adapt to the changes. Darwin explained in naturalistic terms how animals adapt to their environments: traits that contribute to an organism's ability to survive and reproduce in specific environments will become more common. The most “fit”—those organisms best suite
d to the current environmental conditions in which they live—have survived over eons of the history of life on earth to cocreate ecosystems full of animals and plants. Our own bodies are full of evident adaptations: eyes for seeing, ears for hearing, feet for walking on, and so forth.
But what about hands? Feet are adapted to be primarily weight-bearing structures (rather than grasping structures, as in the apes) and that is what we primarily use them for. But we use our hands in many ways: for fine-scale manipulation, greeting, pointing, stimulating a sexual partner, writing, throwing, and cooking, among other uses. So which of these uses express what hands are “for,” when all of them express what hands do?
Gould and Lewontin (1979) illustrate the problem with assuming that the function of a trait defines its evolutionary cause. Consid
er the case of Dr. Pangloss—the protagonistic of Voltaire’s Candide—who believed that we lived in the best of all possible worlds. Gould and Lewontin use his pronouncement that “noses were made for spectacles and so we have spectacles” to demonstrate the problem with assuming any trait has evolved for a specific purpose. Identifying a function of a trait does not necessitate that the function is the ultimate cause of the trait. Individual traits are not under selection pressures in isolation; in fact, an entire organism must be able to survive and reproduce in their environment. When natural selection results in adaptations, changes that occur in some traits can have cascading effects throughout the phenotype and features that are not under selection pressure can also change.
Dig Deeper: Rapid Mitochondrial Evolution in Stingless Bees
A striking example of this interactional evolutionary change comes from recent research on stingless bees. When observing the mitochondrial genome (mitogenome) in two Australian stringless bees in the genus Tetragonula—T. carbonaria and T. hockingsi—they exhibit a rare duplication of the entire mitogenome and show rapid divergence from other members of their species (Françoso et al., 2023). This accelerated evolution is hypothesized to result from factors such as low effective population, founder effects, and genome duplication triggered by environmental stressors. This phenomenon echoes the earlier work by Conrad H. Waddington (1956) mentioned in this chapter, whose experiments exposing fruit fly embryos to ether induced the development of additional wings and thoraces, changes that later became heritable under stable conditions (Shook et al., 2023b). Both cases highlight how organisms can respond to intense environmental pressures through dramatic developmental and genetic shifts.
The mitochondria genetics influence the energy synthesis of the cells and in most animals, the mitogenome remains relatively stable (Shook et al., 2023a); however, Tetragonula species appear to possess an unusual capacity for rapid sequence rearrangement and complete genome duplication, suggesting that their mitogenomes play an important adaptive role. Comparing these genomes with other species such as Lepidotrigona—which shows rearrangements but no duplication— provides a unique opportunity to examine how different lineages respond to similar ecological pressures. Françoso et al. (2023) found that Tetragonula mitogenomes form amphimeric circular structures in which two complete genomes are joined head-to-tail, an extremely rare configuration. These arrangements, including inversions and translocations of gene blocks such as ND6, CytB, ND1, and several rRNA and tRNA genes, are far less common in other bee genera. This pattern supports the idea proposed by Gould & Eldredge (1977) that species are fundamentally unstable entities subject to bursts of rapid change in response to environmental pressures, rather than progressing along a single linear pathway. It is important to note that not all species within the genus exhibit the same degree or type of genomic flexibility. While T. carbonaria and T. hockingsi show full mitogenome duplications, the aforementioned Lepidotrigona species show only partial rearrangements despite facing similar environmental conditions. This variation challenges deterministic assumptions that evolution necessarily moves species toward optimal forms. Instead, it illustrates that evolution often involves trial-and-error shifts shaped by constraint, chance, and ecological stress.
Although further research is needed to determine precisely what triggers such rapid genomic events, the evidence demonstrates that mitochondria play an active role in shaping evolutionary pathways. These findings complicate traditional gradualist models and highlight the importance of examining molecular responses to environmental pressures.
There is an important lesson in recognizing that what things do in the present is not a good guide to understanding why they came to exist. Gunpowder was invented for entertainment—only later was it adopted for killing people. The Internet was invented to decentralize computers in case of a nuclear attack—and only later adopted for social media. Apes have short thumbs and use their hands in locomotion; our ancestors stopped using their hands in locomotion by about six million years ago and had fairly modern-looking hands by about two million years ago. We can speculate that a combination of selection for abstract thought and dexterity led to evolution of the human hand, with its capability for toolmaking that exceeds what apes can do (see Figure 3.5). But let’s face it—how many tools have you made today?
Consequently, we are obliged to see the human foot as having a purpose to which it is adapted and the human hand as having multiple purposes, most of which are different from what it originally evolved for. Paleontologists Gould and Elisabeth Vrba suggested that an original use be regarded as an adaptation and any additional uses be called “exaptations.” Thus, we would consider the human hand to be an adaptation for toolmaking and an exaptation for writing. So how do we know whether any particular feature is an adaptation, like the walking foot, rather than an exaptation, like the writing hand? Or more broadly, how can we reason rigorously from what a feature does to what it evolved for?
The answer to the question “what did this feature evolve for?” creates an origin myth. This origin myth contains three assumptions: (1) features can be isolated as evolutionary units; (2) there is a specific reason for the existence of any particular feature; and (3) there is a clear and simplistic explanation for why the feature evolved.
The first assumption was appreciated a century ago as the “unit-character problem.” Are the units by which the body grows and evolves the same as units we name? This is clearly not the case: we have genes and we have noses, and we have genes that affect noses, but we don’t have “nose genes.” What is the relationship between the evolving elements that we see, identify, and name, and the elements that biologically exist and evolve? It is hard to know, but we can use the history of science as a guide to see how that fallacy has been used by earlier generations. Back in the 19th century, the early anatomists argued that since the brain contained the mind, they could map different mental states (acquisitiveness, punctuality, sensitivity) onto parts of the brain. Someone who was very introspective, say, would have an enlarged introspection part of the brain, a cranial bulge to represent the hyperactivity of this mental state. The anatomical science was known as phrenology, and it was predicated on the false assumption that units of thought or personality or behavior could be mapped to distinct parts of the brain and physically observed (see Figure 3.6). This is the fallacy of reification, imagining that something named is something real.

The second assumption, that everything has a reason, has long been recognized as a core belief of religion. Our desire to impose order and simplicity on the workings of the universe, however, does not constrain it to obey simple and orderly causes. Magic, witchcraft, spirits, and divine agency are all powerful explanations for why things happen. Consequently, it is probably not a good idea to lump natural selection in with those. Sometimes things do happen for a reason, of course, but other times things happen as byproducts of other things, or for very complicated and entangled reasons, or for no reason at all. What phenomena have reasons and thereby merit explanation? Chimpanzees have very large testicles, and we think we know why: their promiscuous sexual behavior triggers intense competition for high sperm count. But chimpanzees also have very large ears, but much less scientific attention has been paid to this trait (see Figure 3.7). Why not? Why should there be a reason for chimp testicles but not for chimp ears? What determines the kinds of features that we try to explain, as opposed to the ones that we do not? Again, the assumption that any specific feature has a reason is metaphysical; that is to say, it may be true in any particular case, but to assume it in all cases is gratuitous.
And third, the possibility of knowing what the reason for any particular feature is, assuming that it has one, is a challenge for evolutionary epistemology (the theory of how we know things). Consider the big adaptations of our lineage: bipedalism and language. Nobody doubts that they are good, and they evolved by natural selection, and we know how they work. But why did they evolve? If talking and walking are simply better than not talking and not walking, then why did they evolve in just a single branch of the ape lineage in the primate family tree? We don’t know what bipedalism evolved for, although there are plenty of speculations: walking long distances, running long distances, cooling the head, seeing over tall grass, carrying babies, carrying food, wading, threatening, counting calories, sexual display, and so on. Neither do we know what language evolved for, although there are speculations: coordinating hunting, gossiping, manipulating others. But it is also possible that bipedality is simply the way that a small arboreal ape travels on the ground, if it isn’t in the treetops. Or that language is simply the way that a primate with small canine teeth and certain mental propensities comes to communicate. If that were true, then there might be no reason for bipedality or language: having the unique suite of preconditions and a fortuitous set of circumstances simply set them in motion, and natural selection elaborated and explored their potentials. It is possible that walking and talking simply solved problems that no other lineage had ever solved; but even if so, the fact remains that the rest of the species in the history of life have done pretty well without having solved them.
It is certainly very optimistic to think that all three assumptions (that organisms can be meaningfully atomized, that everything has a reason, and that we can know the reason) would be simultaneously in effect. Indeed, just as there are many ways of adapting (genetically, epigenetically, behaviorally, culturally), there are also many ways of being nonadaptive, which would imply that there is no reason at all for the feature in question.
First, there is the element of randomness of population histories. There are more cases of sickle-cell anemia among sub-Saharan Africans than other peoples, and there is a reason for it: carriers of sickle-cell anemia have a resistance to malaria, which is more frequent in parts of Africa (as discussed in Chapters 4 and 14). But there are more cases of a blood disease called variegated porphyria, a rare genetic metabolic disorder, in the Afrikaners of South Africa (descendants of mostly Dutch settlers in the 17th century) than in other peoples, and there is no reason for it. Yet we know the cause: One of the founding Dutch colonial settlers had the allele–a variant of a gene–and everyone in South Africa with it today is her descendant. But that is not a reason—that is simply an accident of history.
Second, there is the potential mismatch between the past and the present. The value of a particular feature in the past may be changed as the environmental circumstances change. Our species is diurnal, and our ancestors were diurnal. But beginning around a few hundred thousand years ago, our ancestors could build fires, which extended the light period, which was subsequently further amplified by lamps and candles. And over the course of the 20th century, electrical power has made it possible for people to stay up very late when it is dark—working, partying, worrying—to a greater extent than any other closely related species. In other words, we evolved to be diurnal, yet we are now far more nocturnal than any of our recent ancestors or close relatives. Are we adapting to nocturnality? If so, why? Does it even make any sense to speak of the human occupation of a nocturnal ape niche, despite the fact that we empirically seem to be doing just that? And if so, does it make sense to ask what the reason for it is?
Third, there is a genetic phenomenon known as a selective sweep, or the hitchhiker effect. Imagine three genes—A, B, and C—located very closely together on a chromosome. They each have several variants, or alleles, in the population. Now, for whatever reason, it becomes beneficial to have one of the B alleles, say B4; this B4 allele is now under strong positive selection. Obviously, we will expect future generations to be characterized by mostly B4. But what was B4 attached to? Because whatever A and C alleles were adjacent to it will also be quickly spread, simply by virtue of the selection for B4. Even if the A and C alleles are not very good, they will spread because of the good B4 allele between them. Eventually the linkage groups will break up because of genetic crossing-over in future generations. But in the meantime, some random version of genes A and C are proliferating in the species simply because they are joined to superior allele B4. And clearly, the A and C alleles are there because of selection—but not because of selection for them!
Fourth, some features are simply consequences of other properties rather than adaptations to external conditions. We already noted the phenomenon of allometric growth, in which some physical features have to outgrow others to maintain function at an increased size. Can we ask the reason for the massive brow ridges of Homo erectus, or are brow ridges simply what you get when you have a conjunction of thick skull bones, a large face, and a sloping forehead—and, thus, again would have a cause but no reason?
Fifth, some features may be underutilized and on the way out. What is the reason for our two outer toes? They aren’t propulsive, they don’t do anything, and sometimes they’re just in the way. Obviously they are there because we are descended from ancestors with five digits on their hands and feet. Is it possible that a million years from now, we will just have our three largest toes, just as the ancestors of the horse lost their digits in favor of a single hoof per limb? Or will our outer toes find another use, such as stabilizing the landings in our personal jet-packs? For the time being, we can just recognize vestigiality as another nonadaptive explanation for the presence of a given feature.
Finally, Darwin himself recognized that many obvious features do not help an animal survive. Some things may instead help an animal breed. The peacock’s tail feathers do not help it eat, but they do help it mate. There is competition, but only against half of the species. Darwin called this sexual selection. Its result is not a fit to the environment but, rather, a fit to the opposite sex. In some species, that is literally the case, as the male and female genitalia have specific ways of anatomically fitting together. The specific form is less important than the specific match, so inquiring about the reason for a particular form of the reproductive anatomy may be misleading. The specific form may be effectively random, as long as it fits the opposite sex and is different from the anatomies of other species. Nor is sexual selection the only form of selection that can affect the body differently from natural selection. Competition might also take place between biological units other than organisms—perhaps genes, perhaps cells, or populations, or species. The spread of cultural things, such as head-binding or cheap refined fructose or forced labor, can have significant effects upon bodies, which are also not adaptations produced by natural selection. They are often adaptive physiological responses to stresses but not the products of natural selection.
With so many paths available by which a physical feature might have organically arisen without having been the object of natural selection, it is unwise to assume that any individual trait is an adaptation. And that generalization applies to the best-known, best-studied, and most materially based evolutionary adaptations of our lineage. But our cultural behaviors are also highly adaptive, so what about our most familiar social behaviors? Patriarchy, hierarchy, warfare—are these adaptations? Do they have reasons? Are they good for something?
This is where some sloppy thinking has been troublesome. What would it mean to say that patriarchy evolved by natural selection in the human species? If, on the one hand, it means that the human mind evolved by natural selection to be able to create and survive in many different kinds of social and political regimes, of which patriarchy is one, then biological anthropologists will readily agree. If, on the other hand, it means that patriarchy evolved by natural selection, that implies that patriarchy is genetically determined (since natural selection is a genetic process) and out-reproduced the alleles for other, more egalitarian, social forms. This in turn would imply that patriarchy is an adaptation and therefore of some beneficial value in the past and has become an ingrained part of human nature today. This would be bad news, say, if you harbored ambitions of dismantling it. Dismantling patriarchy in that case would be to go against nature, a futile gesture. In other words, this latter interpretation would be a naturalistic manifesto for a conservative political platform: don’t try to dismantle the patriarchy, because it is within us, the product of evolution—suck it up and live with it.
Here, evolution is being used as a political instrument for transforming the human genome into an imaginary glass ceiling against equality. There is thus a convergence between the pseudo-biology of crude adaptationism (the idea that everything is the product of natural selection) and the pseudo-biology of hereditarianism. Naturalizing inequality is not the business of evolutionary theory, and it represents a difficult moral position for a scientist to adopt, as well as a poor scientific position.
Evolution of the Anthropocene

Under the previously explored Adaptationism and Panglossian Paradigm, it is explained that human evolution is constantly occurring even throughout periods of ecological stability. While this acknowledges evolution as an ongoing process of change, it fails to explore the implications of such on the alteration of other species and ecosystems.
The emergence of the Anthropocene, driven by human activity, though not recognized as an official epoch, is seen as a transformative event comparable to other major historical shifts such as the Ordovician Biodiversification (UNESCO, 2024). Given its scale, it is crucial to inform scholars about the impact of our social and cultural evolution on the rest of the world. Richard Robbins’ Global Problems and Culture of Capitalism explains how the modern culture of consumption has been extremely successful at accommodating populations of people far larger than previously possible. Robbins claims that the globalization attributed to capitalism has allowed the world to make full use of its environmental resources, providing necessities and innovative technologies to humans all over the world (Robbins & Dowty, 2019). In other words, capitalism is an anthropocentric cultural system that highly benefits humans and facilitates our survival with little regard to the development and survival of other forms of life. It would be highly relevant to introduce the idea that our cultural evolution and capacity to modify the environment to meet our needs have established new environmental conditions in which the human species' survival and reproduction rate expand at the detriment of ecosystems and endangerment of other primates and non-human species.
According to the International Union for Conservation of Nature’s Red List of Threatened Species, there are currently over 169,000 species listed, with more than 47,000 species at risk of extinction — including 41% of amphibians, 26% of mammals, 26% of freshwater fishes, 12% of birds, and many others (IUCN, 2025). Human lifestyles are causing changes that—if not taken into consideration—could lead to our extinction as a species. The recognition that our evolutionary behavioural development is causing environmental destruction may be the first step for our species to take accountability for the damage that it is causing to others and prevent further damage.
Summary
Now that you have finished reading this chapter, you are equipped to understand the historical and political dimensions of evolution. Evolution is an ongoing process of change and diversification. Evolutionary theory is a tool that we use to understand this process. The development of evolutionary theory is shaped both by scientific innovation and political engagement. Since Darwin first articulated natural selection as an observable mechanism by which species adapt to their environments, our understanding of evolution has grown. Initially, scientists focused on the adaptive aspects of evolution. However, with the emergence of genetics, our understanding of heredity and the level at which evolution acts has changed. Genetics led to a focus on the molecular dimensions of evolution. For some, this focus resulted in reductive accounts of evolution. Further developments in our understanding of evolution shifted our view to epigenetic processes and how organisms shape their own evolutionary pressures (e.g., niche construction).
Evolutionary theory will continue to develop in the future as we invent new technologies, describe new dimensions of biology, and experience cultural changes. Current innovations in evolutionary theory are asking us to consider evolutionary forces beyond natural selection and genetics to include the ways organisms shape their environments (niche construction), inheritances beyond genetics (inclusive inheritance), constraints on evolutionary change (developmental bias), and the ability of bodies to change in response to external factors (plasticity). The future of evolutionary theory looks bright as we continue to explore these and other dimensions. Biological anthropology is well-positioned to be a lively part of this conversation, as it extends standard evolutionary theory by considering the role of culture, social learning, and human intentionality in shaping the evolutionary trajectories of humans (Zeder 2018). Remember, at root, human evolutionary theory consists of two propositions: (1) the human species is descended from other similar species and (2) natural selection has been the primary agent of biological adaptation. Pretty much everything else is subject to some degree of contestation.
Review Questions
- How is the study of your ancestors biopolitical, not just biological? Does that make it less scientific or differently scientific?
- What was gained by reducing organisms to genotypes and species to gene pools? What is gained by reintroducing bodies and species into evolutionary studies?
- How do genetic or molecular studies complement anatomical studies of evolution?
- How are you reducible to your ancestry? If you could meet your ancestors from the year 1700 (and you would have well over a thousand of them!), would their lives be meaningfully similar to yours? Would you even be able to communicate with them?
- The molecular biologist François Jacob argued that evolution is more like a tinkerer than an engineer. In what ways do we seem like precisely engineered machinery, and in what ways do we seem like jerry-rigged or improvised contraptions?
- How might biological anthropology contribute to future developments in evolutionary theory?
Key Terms
Adaptation: A fit between the organism and environment.
Adaptationism: The idea that everything is the product of natural selection.
Allele: A genetic variant.
Allometry: The differential growth of body parts.
Canalization: The tendency of a growing organism to be buffered toward normal development.
Epigenetics: The study of how genetically identical cells and organisms (with the same DNA base sequence) can nevertheless differ in stably inherited ways.
Eugenics: An idea that was popular in the 1920s that society should be improved by breeding “better” kinds of people.
Evo-devo: The study of the origin of form; a contraction of “evolutionary developmental biology.”
Exaptation: An additional beneficial use for a biological feature.
Extinction: The loss of a species from the face of the earth.
Gene: A stretch of DNA with an identifiable function (sometimes broadened to include any DNA with recognizable structural features as well).
Gene pool: Hypothetical summation of the entire genetic composition of population or species.
Genotype: Genetic constitution of an individual organism.
Hereditarianism: The idea that genes or ancestry is the most crucial or salient element in a human life. Generally associated with an argument for natural inequality on pseudo-genetic grounds.
Hox genes: A group of related genes that control for the body plan of an embryo along the head-tail axis.
Inheritance of acquired characteristics: The idea that you pass on the features that developed during your lifetime, not just your genes; also known as Lamarckian inheritance.
Natural selection: A consistent bias in survival and fertility, leading to the overrepresentation of certain features in future generations and an improved fit between an average member of the population and the environment.
Niche construction: The active engagement by which species transform their surroundings in favorable ways, rather than just passively inhabiting them.
Phenotype: Observable manifestation of a genetic constitution, expressed in a particular set of circumstances. The suite of traits of an organism.
Phrenology: The 19th-century anatomical study of bumps on the head as an indication of personality and mental abilities.
Plasticity: The tendency of a growing organism to react developmentally to its particular conditions of life.
Punctuated equilibria: The idea that species are stable through time and are formed very rapidly relative to their duration. (The opposite theory, that species are unstable and constantly changing through time, is called phyletic gradualism.)
Scientific racism: The use of pseudoscientific evidence to support or legitimize racial hierarchy and inequality.
Sexual selection: Natural selection arising through preference by one sex for certain characteristics in individuals of the other sex.
Species selection: A postulated evolutionary process in which selection acts on an entire species population, rather than individuals.
For Further Exploration
Ackermann, Rebecca Rogers, Alex Mackay, and Michael L. Arnold. 2016. “The Hybrid Origin of ‘Modern’ Humans.” Evolutionary Biology 43 (1): 1–11.
Bateson, Patrick, and Peter Gluckman. 2011. Plasticity, Robustness, Development and Evolution. New York: Cambridge University Press.
Cosans, Christopher E. 2009. Owen's Ape and Darwin's Bulldog: Beyond Darwinism and Creationism. Bloomington, IN: Indiana University Press.
Desmond, Adrian, and James Moore. 2009. Darwin's Sacred Cause: How a Hatred of Slavery Shaped Darwin's Views on Human Evolution. New York: Houghton Mifflin Harcourt.
Dobzhansky, Theodosius, Francisco J. Ayala, G. Ledyard Stebbins, and James W. Valentine. 1977. Evolution. San Francisco: W.H. Freeman and Company.
Fuentes, Agustín. 2017. The Creative Spark: How Imagination Made Humans Exceptional. New York: Dutton.
Haraway, Donna J. 1989. Primate Visions: Gender, Race, and Nature in the World of Modern Science. New York: Routledge.
Huxley, Thomas. 1863. Evidence as to Man's Place in Nature. London: Williams & Norgate.
Jablonka, Eva, and Marion J. Lamb. 2005. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life. Cambridge, MA: The MIT Press.
Kuklick, Henrika, ed. 2008. A New History of Anthropology. New York: Blackwell.
Laland, Kevin N., Tobias Uller, Marcus W. Feldman, Kim Sterelny, Gerd B. Muller, Armin Moczek, Eva Jablonka, and John Odling-Smee. 2015. “The Extended Evolutionary Synthesis: Its Structure, Assumptions and Predictions.” Proceedings of the Royal Society, Series B 282 (1813): 20151019.
Lamarck, Jean Baptiste. 1809. Philosophie Zoologique. Paris: Dentu.
Landau, Misia. 1991. Narratives of Human Evolution. New Haven: Yale University Press.
Lee, Sang-Hee. 2017. Close Encounters with Humankind: A Paleoanthropologist Investigates Our Evolving Species. New York: W. W. Norton.
Livingstone, David N. 2008. Adam's Ancestors: Race, Religion, and the Politics of Human Origins. Baltimore: Johns Hopkins University Press.
Marks, Jonathan. 2015. Tales of the Ex-Apes: How We Think about Human Evolution. Berkeley, CA: University of California Press.
Pigliucci, Massimo. 2009. “The Year in Evolutionary Biology 2009: An Extended Synthesis for Evolutionary Biology.” Annals of the New York Academy of Sciences 1168: 218–228.
Simpson, George Gaylord. 1949. The Meaning of Evolution: A Study of the History of Life and of Its Significance for Man. New Haven: Yale University Press.
Sommer, Marianne. 2016. History Within: The Science, Culture, and Politics of Bones, Organisms, and Molecules. Chicago: University of Chicago Press.
Stoczkowski, Wiktor. 2002. Explaining Human Origins: Myth, Imagination and Conjecture. New York: Cambridge University Press.
Tattersall, Ian, and Rob DeSalle. 2019. The Accidental Homo sapiens: Genetics, Behavior, and Free Will. New York: Pegasus.
References
Barton, Robert A. 1996. "Neocortex Size and Behavioural Ecology in Primates." Proceedings of the Royal Society of London. Series B: Biological Sciences 263 (1367): 173–177.
Bodmer, Walter, and Robin McKie. 1997. The Book of Man: The Hman Genome Project and the Quest to Discover our Genetic Heritage. Oxford University Press.
Brassard, A., Perron-Laplante, J., Lachapelle, É., Pierrepont, C. de, & Péloquin, K. (2018). Oversexualization among emerging adults: Preliminary associations with romantic attachment and intimacy. The Canadian Journal of Human Sexuality, 27(3), 235–247. Project Muse.
Carroll, S. B. (2005). Evolution at Two Levels: On Genes and Form. PLoS Biology, 3(7). Public Library of Science. https://doi.org/10.1371/journal.pbio.0030245
Chudek, M., Muthukrishna, M., & Henrich, J. (2015). Cultural evolution. The Handbook of Evolutionary Psychology, 1–21. https://doi.org/10.1002/9781119125563.evpsych230
Darwin, Charles. 1859. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life. London: J. Murray.
Darwin, Charles. 1871. The Descent of Man, and Selection in Relation to Sex. London: J. Murray.
Dawkins, Richard. 1976. The Selfish Gene. Oxford University Press.
Deacon, T. W. 1998. The Symbolic Species: The Co-evolution of Language and the Brain. W. W. Norton & Company.
Eldredge, N., and S. J. Gould. 1972. "Punctuated Equilibria: An Alternative to Phyletic Gradualism." In Models in Paleobiology, edited by T. J. Schopf, 82–115. San Francisco: W. H. Freeman.
Françoso, E., Zuntini, A. R., Ricardo, P. C., Santos, P. K. F., de Souza Araujo, N., Silva, J. P. N., Gonçalves, L. T., Brito, R., Gloag, R., Taylor, B. A., Harpur, B. A., Oldroyd, B. P., Brown, M. J. F., & Arias, M. C. (2023). Rapid evolution, rearrangements and whole mitogenome duplication in the Australian stingless bees Tetragonula (Hymenoptera: Apidae): A steppingstone towards understanding mitochondrial function and evolution. International Journal of Biological Macromolecules, 242. Elsevier. https://doi.org/10.1016/j.ijbiomac.2023.124568
Gayon, J. (2010). Sexual selection: Another Darwinian process. Comptes Rendus Biologies, 333(2), 134–144. MEDLINE. https://doi.org/10.1016/j.crvi.2009.12.001
Goldman, E. A., & Sterner, K. N. (2023). Environment, Epigenetics, and the Pace of Human Aging. Annual Review of Anthropology, 52, 279–294. Annual Reviews. https://doi.org/10.1146/annurev-anthro-052721-090516
Gould, S. J., & Eldredge, N. (1977). Punctuated Equilibria: The Tempo and Mode of Evolution Reconsidered. Paleobiology, 3(2), 115–151. JSTOR Biological Sciences Collection.
Gould, Stephen J. 2003. The Structure of Evolutionary Theory. Cambridge, MA: Harvard University Press.
Gould, Stephen J. 1996. Mismeasure of Man. New York: WW Norton & Company.
Gould, Stephen Jay, and Richard C. Lewontin. 1979. "The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme." Proceedings of the Royal Society of London. Series B: Biological Sciences 205 (1151): 581–598.
Haeckel, Ernst. 1868. Natürliche Schöpfungsgeschichte. Berlin: Reimer.
Huxley, Thomas Henry. 1863. Evidence as to Man’s Place in Nature. London: Williams and Norgate.
IUCN. 2025. The IUCN Red List of Threatened Species. Version 2025-1. https://www.iucnredlist.org. Accessed on 30 July 2025.
Kaufman, Thomas C., Mark A. Seeger, and Gary Olsen. 1990. "Molecular and Genetic Organization of the Antennapedia Gene Complex of Drosophila melanogaster." Advances in Genetics 27: 309–362.
Kellogg, Vernon. 1917. Headquarters Nights. Boston: The Atlantic Monthly Press.
Kevles, Daniel J., and Leroy Hood. 1993. The Code of Codes: Scientific and Social Issues in the Human Genome Project. Cambridge, MA: Harvard University Press.
Lewontin, Richard, Steven Rose, and Leon Kamin. 2017. Not in Our Genes : Biology, Ideology, and Human Nature, 2nd ed. Chicago: Haymarket Books.
Lloyd, Elisabeth A., and Stephen J. Gould. 1993. "Species Selection on Variability." Proceedings of the National Academy of Sciences 90 (2): 595–599.
Marks, Jonathan. 2015. “The Biological Myth of Human Evolution.” In Biologising the Social Sciences: Challenging Darwinian and Neuroscience Explanations, edited by David Canter and David A. Turner, 59–78. London: Routledge.
Meaney, M. J. (2001). Maternal care, gene expression, and the transmission of individual differences in stress reactivity across generations. Annual Review of Neuroscience, 24, 1161–1192. MEDLINE.
Monypenny, William Flavelle, and George Earle Buckle. 1929. The Life of Benjamin Disraeli, Earl of Beaconsfield, Volume II: 1860–1881. London: John Murray.
Potts, Rick. 1998. “Variability Selection in Hominid Evolution.” Evolutionary Anthropology 7: 81–96.
Punnett, R. C. 1905. Mendelism. Cambridge: Macmillan and Bowes.
Robbins, R., & Dowty, R. (2019). Robbins Richard, Global problems and culture of capitalism (7th ed.). Pearson.
Shapiro, Robert. 1991. The Human Blueprint: The Race to Unlock the Secrets of Our Genetic Script. New York: St. Martin’s Press.
Schafte, K., & Bruna, S. (2023). The influence of intergenerational trauma on epigenetics and obesity in Indigenous populations—A scoping review. Epigenetics, 18(1), 2260218. MEDLINE. https://doi.org/10.1080/15592294.2023.2260218
Shultz, Susanne, Emma Nelson, and Robin Dunbar. 2012. "Hominin Cognitive Evolution: Identifying Patterns and Processes in the Fossil and Archaeological Record." Philosophical Transactions of the Royal Society B: Biological Sciences 367 (1599): 2130–2140.
Shook, B., Nelson, K., Braff, L., & Aguilera, K. (2023a). Molecular Biology and Genetics. https://opentextbooks.concordia.ca/explorationsversiontwo/chapter/3/
Shook, B., Nelson, K., Braff, L., & Aguilera, K. (2023b). Social and Biopolitical Dimensions of Evolutionary Thinking. https://opentextbooks.concordia.ca/explorationsversiontwo/chapter/social-and-bi opolitical-dimensions-of-evolutionary-thinking/
Spencer, Herbert. 1864. Principles of Biology. London: Williams and Norgate.
UNESCO. (2024). The Anthropocene. International Union of Geological Sciences. https://www.iugs.org/_files/ugd/f1fc07_40d1a7ed58de458c9f8f24de5e739663.pdf?index=true
Vicedo, M. (2025). Charlotte Perkins Gilman: A Pragmatist Framework for Constructing a New Humanhood. Journal of the History of the Behavioral Sciences, 61(4), e70041. MEDLINE. https://doi.org/10.1002/jhbs.70041
Watson, James D. 1990. "The Human Genome Project: Past, Present, and Future." Science 248 (4951): 44–49.
Wray, G. A. (2007). The evolutionary significance of cis-regulatory mutations. Nature Reviews. Genetics, 8(3), 206–216. MEDLINE.
Waddington CH. Genetic assimilation of the bithorax phenotype. Evolution. 1956;10:1–13. doi: 10.1111/j.1558-5646.1956.tb02824.x.
Yengo, L., Vedantam, S., Marouli, E., Sidorenko, J., Bartell, E., Sakaue, S., Graff, M., Eliasen, A.U., Jiang, Y., Raghavan, S. and Miao, J., 2022. A saturated map of common genetic variants associated with human height. Nature, 610 (7933): 704-712.
Zeder, Melinda A. 2018. "Why Evolutionary Biology Needs Anthropology: Evaluating Core Assumptions of the Extended Evolutionary Synthesis." Evolutionary Anthropology: Issues, News, and Reviews 27 (6): 267–284.

{kind=link}
{kind=link}
,_homme_de_N%C3%A9andertal.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hardy_Michel_bpt6k5567846s_(2).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}