2 A History of Evolutionary Thought
Joylin Namie, Ph.D., Truckee Meadows Community College
Learning Objectives
- Identify and describe the major developments in scientific thought that led to the discovery of evolutionary processes.
- Explain how natural selection works and results in evolutionary change over time.
- Explain what is meant by the “Modern Synthesis” and its impacts on evolutionary thought.
- Discuss the teaching of human evolution in the U.S. and abroad.
The Beginnings of Evolutionary Thinking
Throughout our evolutionary history, humans have developed an understanding of the natural world as they interacted with and extracted resources from it. To survive, our earliest ancestors possessed an understanding of the physical environment, including weather patterns, animal behavior, edible and medicinal plants, locations of water, and seasonal cycles. Many ancient cultures, including those of the Americas (Dunbar-Ortiz 2014), Mesopotamia, and Egypt, left writings, hieroglyphics, and stories passed down through oral tradition detailing their understanding of the natural environment, human and zoological anatomy, botany, and medical practices (Moore 1993).
There are also over 2,000 years of organized thought and writing regarding evolution, including contributions from Greek, Roman, and Islamic scholars. Three examples of note are included here. The Greek philosopher Aristotle (384–322 BCE) studied the natural world, publishing several volumes on animals based on systematic observations, rather than attributing what he observed to divine intervention, as his contemporaries were doing (Figure 2.1). Aristotle’s system for the biological classification of nearly 500 species of animals was based on his own observations and dissections, interviews with specialists such as beekeepers and fishermen, and accounts of travelers. His nine book History of Animals, published in the 4th century BC (n.d.), was one of the first zoological taxonomies ever created. Aristotle’s primary contribution to the classification of biological species was to recognize that natural groups are based on structure, physiology, mode of reproduction, and behavior (Moore 1993, 39).



Aristotle’s History of Animals also placed animals in a hierarchy, ranking animals above plants due to what he claimed were their abilities to sense the world around them and to move. He also graded animals according to their modes of reproduction. Those giving birth to live young were placed above those who laid eggs. Warm-blooded animals ranked above invertebrates. This concept of “higher” and “lower” organisms was expanded upon by scholars in the Medieval period to form the Scala Naturae (Latin for “ladder of being”). This “Great Chain of Being,” depicting a hierarchy of beings with God at the top and minerals at the bottom (Figure 2.2), was thought by medieval Christians to have been decreed by God; in this Great Chain, humans were placed closer to God than other species. Aristotle’s works were rediscovered by Islamic scholars in the ninth century and translated into Arabic, Syriac, Persian, and later into Latin, becoming part of university curriculum in 13th-century Europe (Lindberg 1992), allowing Aristotle’s works and ideas to influence other thinkers for 2,000 years.

Science also owes a debt to many Arabic scholars. One such Islamic scholar and writer, who built upon the ideas of Aristotle, was Abū ʿUthman ʿAmr ibn Baḥr al-Kinānī al-Baṣrī / al-Jāḥiẓ, known as Al-Jahiz (776–868 CE), who authored over 200 books (El-Zaher 2018; Figure 2.3). His most well-known work was the seven-volume Kitab al-Hayawan or Book of Animals, in which he described over 350 species in zoological detail. Importantly, Al-Jahiz introduced the idea and mechanisms of biological evolution 1,000 years before Darwin proposed the concept of natural selection in 1859 (Love 2020). For instance, Al-Jahiz wrote about the struggle for existence, the transformation of species over time, and environmental factors that influence the process, all ideas that were later espoused by western European scientists in the 19th century. Al-Jahiz wrote:
Animals engage in a struggle for existing, and for resources, to avoid being eaten, and to breed. Environmental factors influence organisms to develop new characteristics to ensure survival, thus transforming them into new species. Animals that survive to breed can pass on their successful characteristics to their offspring. [Masood 2009]

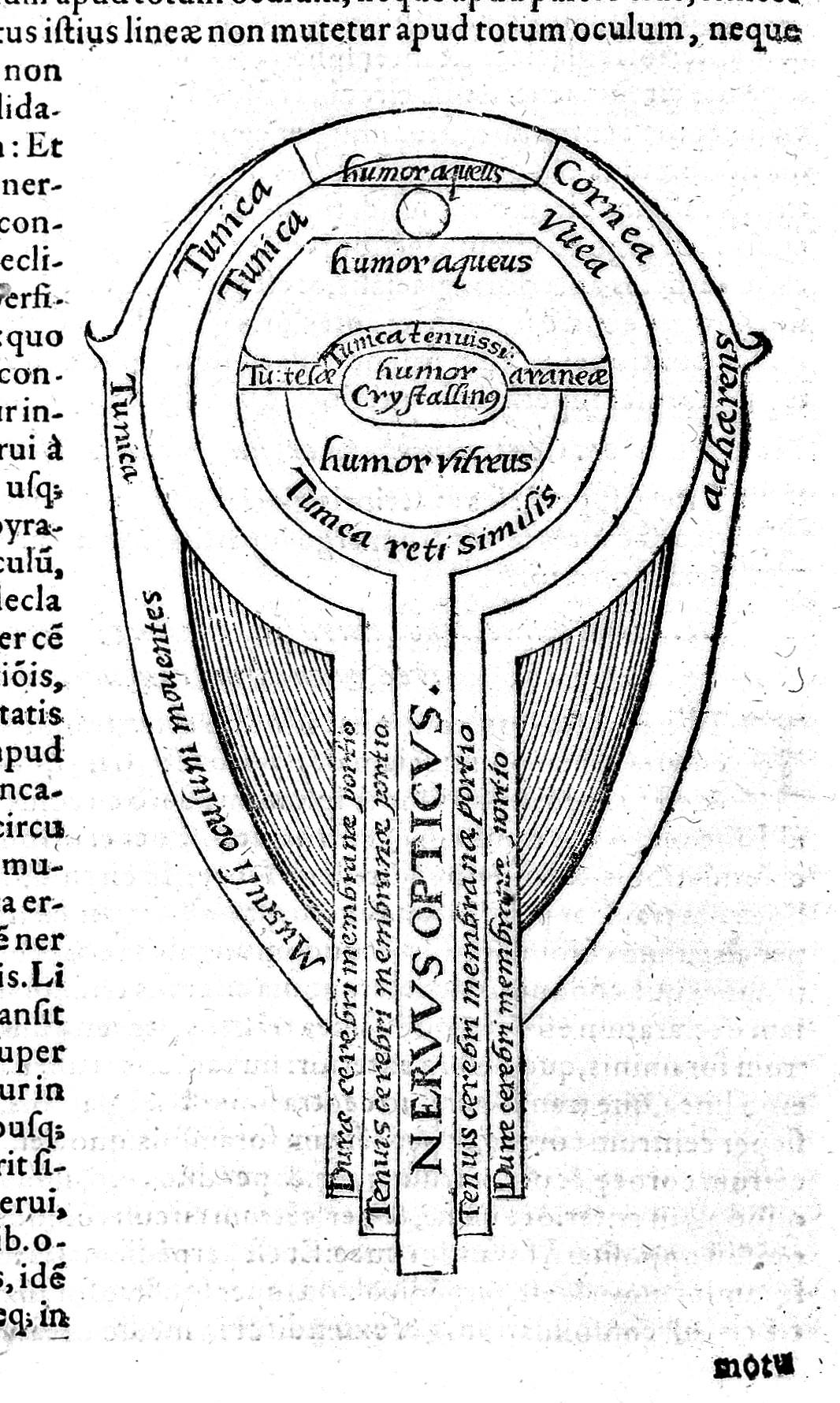
Another important early Islamic scientist is Ibn al-Haytham (965–1040), a 10th-century Islamic scholar who contributed a great deal to our understanding of optics and how human vision works (Figure 2.4; Lindberg 1992, 177–180). Born in what is now Iraq, al-Haytham was a scholar of many disciplines, including mathematics, physics, mechanics, astronomy, philosophy, and medicine. He authored some 200 books in his lifetime and was an expert on Aristotle’s natural philosophy, logic, and metaphysics. In relation to evolution, al-Haytham’s methodology of investigation—specifically, using experiments to verify theory—is similar to what later became known as the modern scientific method. He is most famous for discovering the laws of reflection and refraction over 1,000 years ago and inventing the camera obscura, which was incredibly important for the eventual development of photography. His work is credited for its influence on astronomy, mathematics, and optics, inspiring Galileo, Johannes Kepler, and Sir Isaac Newton (Tasci 2020). As an inspirational scientific figure, al-Haytham was celebrated in 2016 by UNESCO as a trailblazer and the founder of modern optics (Figure 2.5). An International Year of Light was named in his honor and a scholarly conference on his contributions was held to coincide with the 1,000th anniversary of the publication of his Kitāb al-Manāẓir (Book of Optics; UNESCO.org 2015).
The writings of these Islamic scholars as well as similar scientific texts from other cultures were unknown to or unacknowledged by Western scientists until recently. Fortunately, many science teachers are now incorporating this content into their classes (Love 2020).
Western European Evolutionary Thought
Although there have been many different scientific traditions throughout world history, a new global discourse around science emerged in Western Europe in the 19th century. Europeans pointed to the continuing expansion of their colonial power, as well as their military and technological success, as evidence of the efficacy of Western science, which came to dominate on a global scale (Elshakry 2010). The movement toward a global science centered in Western Europe began with formulation of the Scientific Method.
The Scientific Method was first codified by Francis Bacon (1561–1626), an English politician who was likely influenced by the methods of inquiry established by Ibn al-Haytham centuries prior (Tbakhi and Amr 2007). Bacon has been called the founder of empiricism for proposing a system for weighing the truthfulness of knowledge based solely on inductive reasoning and careful observations of natural phenomena. Ironically, he died as a result of trying to scientifically observe the effects of cold on the putrefaction of meat. On a journey out of London, he purchased a chicken and stuffed it with snow for observation, catching a chill in the process. One week later, he died of bronchitis (Urbach, Quinton, and Lea 2023).
The second important development with regard to evolution was the concept of a species. John Ray (1627–1705), an English parson and naturalist, was the first person to publish a biological definition of species in his Historia Plantarum (History of Plants), a three volume work published in 1686, 1688, and 1704. Ray defined a species as a group of morphologically similar organisms arising from a common ancestor. However, we now define a species as a group of similar organisms capable of producing fertile offspring. In keeping with the scientific method, Ray classified plants according to similarities and differences that emerged from observation. He claimed that any seed from the same plant was the same species, even if it had slightly different traits.
The modern period of biological classification began with the work of Carl von Linne (“Carolus Linnaeus”) (1707–1778), a Swedish scientist who laid the foundations for the modern scheme of taxonomy used today. He established the system of binomial nomenclature, in which a species of animal or plant receives a name consisting of two terms: the first term identifies the genus to which it belongs and the second term identifies the species. His original Systema Naturae, published in 1736, went through several editions. By the tenth edition in 1758, mammals incorporated primates, including apes and humans, and the term Homo sapiens was introduced to signify the latter (Paterlini 2007).
Georges-Louis Leclerc, Comte de Buffon (1707–1788), was a prominent French naturalist whose work influenced prominent scientists in the second half of the 18th century. Buffon’s idea that species change over time became a cornerstone of modern evolutionary theory. His technique of comparing similar structures across different species, called comparative anatomy, is still in use today in the study of evolution. He published 36 volumes of Histoire Naturelle during his lifetime and heavily influenced two prominent French thinkers who were to have significant impacts on our understanding of evolution, Georges Cuvier and Jean-Baptiste Lamarck.

Georges Cuvier (1769–1832) was a paleontologist and comparative anatomist (Figure 2.6). One of his first major contributions to the field of evolution was proof that some species had become extinct through detailed and comprehensive analyses of large fossil quadrupeds (Moore 1993, 111). The idea of extinction was not new, but it was challenging to demonstrate if a fossil species was truly extinct or still had living relatives elsewhere. It was also challenging in that it ran counter to religious beliefs of the time. The Bible’s Book of Genesis was interpreted as saying that all species had been created by God in the seven days it took to create the world and that all created species have survived to this day. Extinction was interpreted as implying imperfection, suggesting God’s work was flawed. Also, given that the Earth was calculated to have been created in 4004 B.C.E., based on biblical genealogies, there would not have been enough time for species to disappear (Moore 1993, 112).
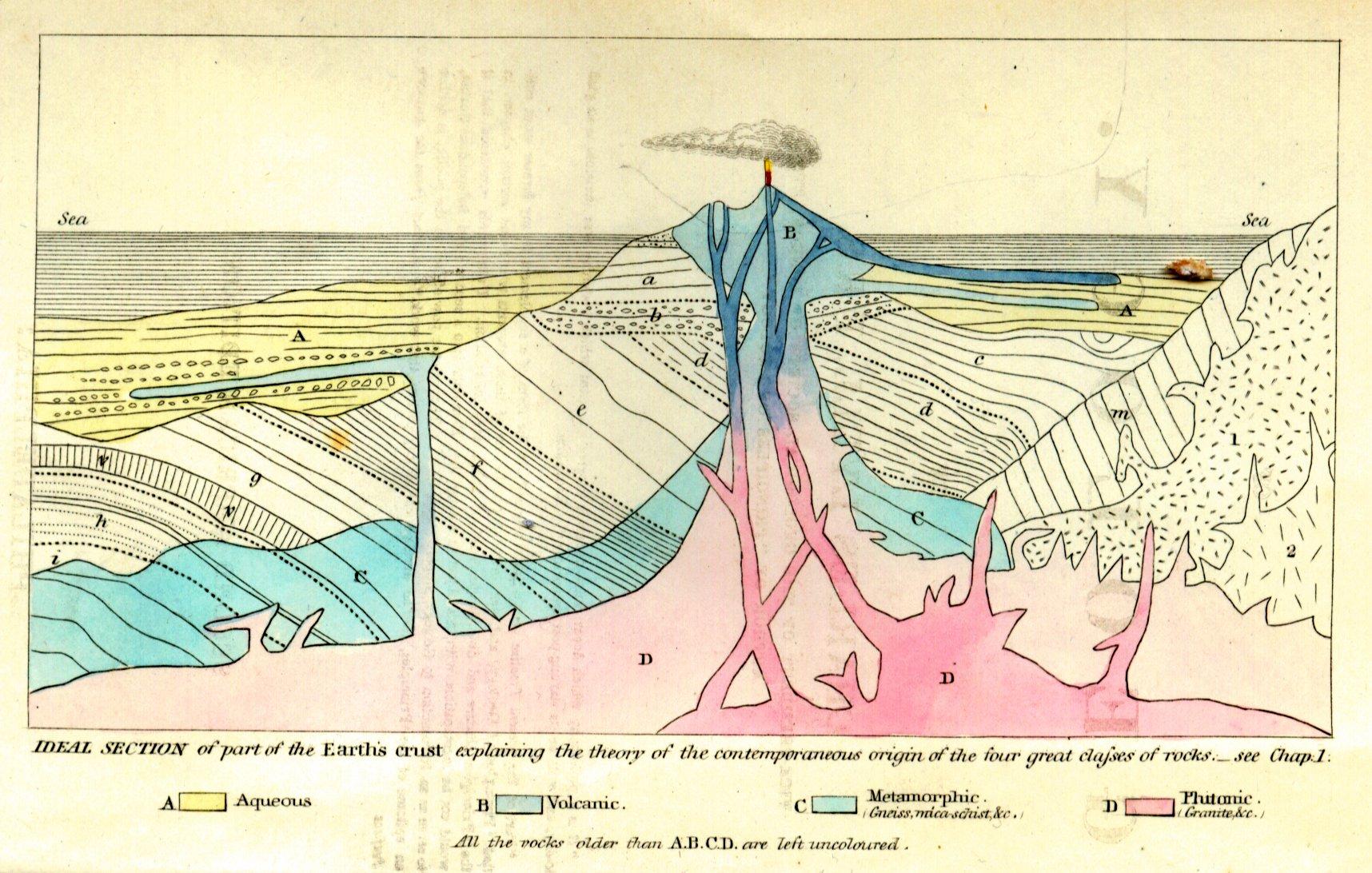
Cuvier was so knowledgeable in this field that he became famous for his ability to reconstruct what an extinct animal looked like from fragmentary remains. He demonstrated that fossil mammoths differed from similar living creatures, such as elephants. His many examples of fossils telling the stories of animals that lived and then disappeared were taken as incontrovertible proof of extinctions (PBS 2001). Where Cuvier went awry was his hypothesis of how extinction worked and its causes. As part of his study of comparative anatomy, Cuvier made observations of stratified layers of rock, or sediment, each containing different species. From this, he drew conclusions that species were “fixed” and did not evolve, but then went extinct, and that different assemblages of fossils occurred at different times in the past, as evidenced by the sedimentary layers (Moore 1993, 118). Cuvier explained this through a theory of catastrophism, which stated that successive catastrophic deluges (akin to Biblical floods) swept over parts of the Earth periodically, exterminating all life. When the waters receded from a particular region, lifeforms from unaffected regions would repopulate the areas that were destroyed, giving rise to a new layer of species that looked different from the layer below it. This theory implied that species were fixed in place and did not evolve and that the Earth was young. In fact, Cuvier postulated that the last catastrophe was a deluge he believed occurred five to six thousand years ago, paving the way for the advent of humans (Moore 1993, 118). Cuvier’s catastrophism became part of an ongoing and vociferous debate between two schools of geology. The catastrophists believed the present state of the earth was the consequence of a series of violent catastrophes of short duration, while the uniformitarians thought it was the result of slow acting geological forces that continue to shape the earth.

James Hutton (1726–1797) was one prominent proponent of uniformitarianism. Based on evidence he found at sites in his native Scotland, Hutton argued that the Earth was much older than previously thought. Examining the geology of Siccar Point, a cliff site on the eastern coast of Scotland (Figure 2.7), Hutton concluded that the intersection of the vertical and horizontal rocks represented a gap in time of many millions of years, during which the lower rocks had been deformed and eroded before the upper layers were deposited on top. From this, Hutton argued sediments are deposited primarily in the oceans, where they become strata, or layers of sedimentary rock. Volcanic action uplifts these strata to form mountains, which are then subject to erosion from rain, rivers, and wind, returning sediment to the oceans (Moore 1993, 121). Hutton’s Theory of the Earth (1788) demanded vast periods of time (known as “deep time”) for such slow-working forces to shape the earth. At the time, he was heavily criticized for this view, as it contradicted the biblical version of the history of creation.
Another Scotsman, who was to become a highly influential geologist and a close friend of Darwin, was Charles Lyell (1797–1875). Lyell was originally a lawyer who began his studies of Geology at Oxford under the tutelage of catastrophist William Buckland, from whom he diverged when Buckland tried to find physical evidence of Noah’s flood from the Christian Bible. Lyell was instead intent on establishing geology as a science based on observation. Building upon Hutton’s ideas (published 50 years earlier), Lyell traveled throughout Europe, documenting evidence of uniformitarianism. During his travels, he cataloged evidence of sea level rise and fall and of volcanoes positioned atop much older rocks. He also found evidence of valleys formed through erosion, mountains resulting from earthquakes, and volcanic eruptions that had been witnessed or documented in the past (University of California Berkeley Museum of Paleontology n.d.). Lyell also espoused the principle that “rocks and strata (layers of rock) increase in age the further down they are in a geological sequence. Barring obvious upheavals or other evidence of disturbance, the same principle must apply to any fossils contained within the rock. The lower down in a sequence of rocks a fossil is, the older it is likely to be (Wood 2005, 12).”
Lyell published the first edition of his three-volume Principles of Geology in 1830–1833 (Figure 2.8). It established geology as a science, underwent constant revisions as new scientific evidence was discovered, and was published in 12 editions during Lyell’s lifetime. In it, he espoused the key concept of uniformitarianism—that “the present is the key to the past.” What this meant was that geological remains from the distant past can be explained by reference to geological processes now in operation and directly observable.
Jean-Baptiste Lamarck (1744–1829) was the first Western scientist to propose a mechanism explaining why and how traits changed in species over time, as well as to recognize the importance of the physical environment in acting on and shaping physical characteristics. Lamarck’s view of how and why species changed through time, known as the “Theory of Inheritance of Acquired Characteristics,” was first presented in the introductory lecture to students in his invertebrate zoology class at the Museum of Natural History in Paris in 1802 (Burkhardt 2013). It was based on the idea that as animals adapted to their environments through the use and disuse of characteristics, their adaptations were passed on to their offspring through reproduction (Figure 2.9). Lamarck was right about the environment having an influence on characteristics of species, as well as about variations being passed on through reproduction. He simply had the mechanism wrong.


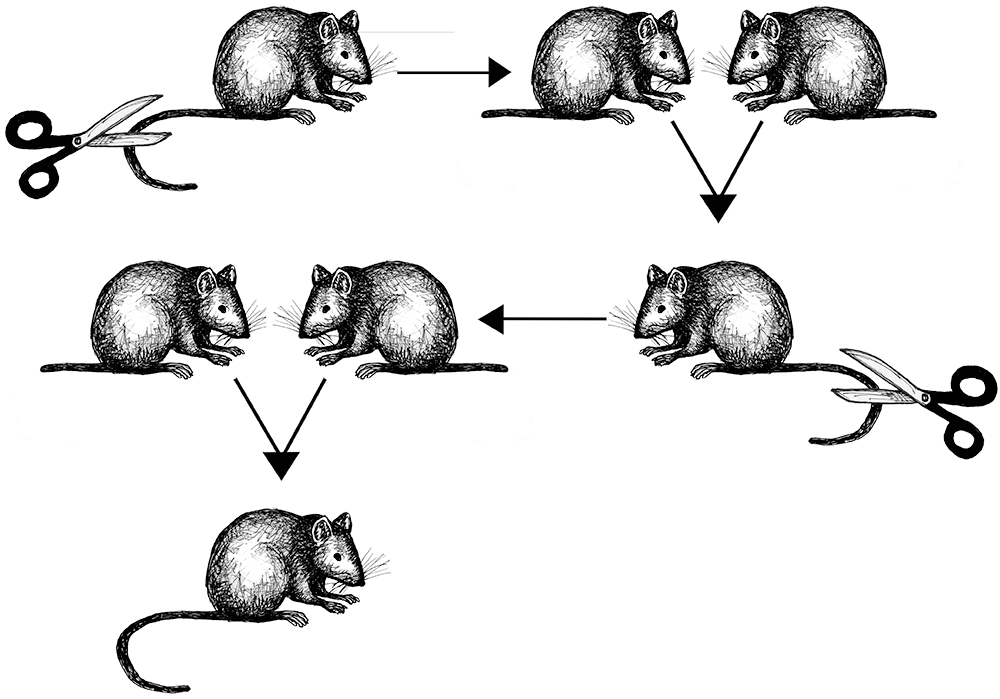
Lamarck’s theory involved a three-step process. Step one involves an animal experiencing a radical change in its environment. Step two is the animal (either individual or species) responding with a new kind of behavior. Step three is how the behavioral change results in morphological (meaning physical) changes to the animal that are successfully passed on to subsequent generations (Ward 2018, 8). Lamarck’s most famous example was the proposition that giraffes actively stretched their necks to reach leaves on tall trees to eat. Over their lifetimes, the continuation of this habit resulted in gradual lengthening of the neck. These longer necks were then passed on to their offspring. Lamarck’s theory was disproved when evolutionary biologist August Weismann published the results of an experiment involving mice (Figure 2.10). Weismann amputated the tails of 68 mice and then successively bred five generations of them, removing the tails of all offspring in each generation, eventually producing 901 mice, all of whom had perfectly healthy long tails in spite of having parents whose tails were missing (Weismann 1889).
How giraffes actually ended up with long necks is a different story. In an environment where the food supply is higher off the ground, and perhaps less available to competing species, giraffes who happened to have slightly longer necks (due to random individual variation and genetic mutation) would be more likely to survive. These giraffes would then be able to reproduce, passing along the slight variation in neck length that would allow their offspring to do the same. Over time, individuals with longer necks would be overrepresented in the population, and neck lengths overall would increase among giraffes. Unfortunately, Lamarck’s ideas challenged the scientific establishment of the time and were rejected. He was discredited and harassed “to the point of loss of money, reputation, and then health” (Ward 2018, 9).
The final piece in the evolutionary puzzle leading up to the theory of natural selection was put forth by Thomas Malthus (1766–1834), who published An Essay on Population in 1798. Malthus lived in England during the time of the Industrial Revolution. It was a time of great poverty and misery when many people migrated from the countryside to squalid, disease-ridden cities to work extremely long hours in dangerous conditions in factories, coal mines, and other industrial workplaces. Birth rates were high and starvation and disease were rampant. Malthus struggled to explain why. His answer was basically the idea of carrying capacity, an ecological concept still in use today. Malthus suggested the rate of population growth exceeded the rate of increase of the human food supply. In other words, people were outgrowing the available food crops. He also suggested that populations of animals and plants were naturally constrained by the food supply, resulting in reductions in population in times of scarcity, “restraining them within the prescribed bounds” (Moore 1993, 147). But, despite significant challenges, some individuals always survived. This was the key to later understandings of evolutionary change in species over time.
The Journey to Natural Selection
In Western European thought, the individual most closely associated with evolution is Charles Darwin (1809–1882). However, as one can see from the individuals and ideas presented in the prior section, he was not the first person to explore evolution and how it might work. In fact, Darwin built upon and synthesized many of the ideas—from geology to biology, ecology, and economy—discussed above. He was simply in the right place at the right time. If he had not worked out his ideas when he did, someone else would have. As a matter of fact, as noted below, someone else did, forcing Darwin to publicly reveal his theory.
Darwin continued his observations and experiments during his formal education, culminating in his graduation from Cambridge in 1831, at which point he was invited to become a gentleman naturalist for a British Royal Navy surveying mission of the globe aboard the H.M.S. Beagle. It is worth noting that Darwin was only 22 years old and the captain’s third choice for the position (Costa 2017), but he proved extremely curious and methodical. The mission departed in December of 1831 and returned five years later (Figure 2.11). During this time, Darwin produced copious notebooks, observations, drawings, and reflections on the natural phenomena he encountered and the experiments he performed.
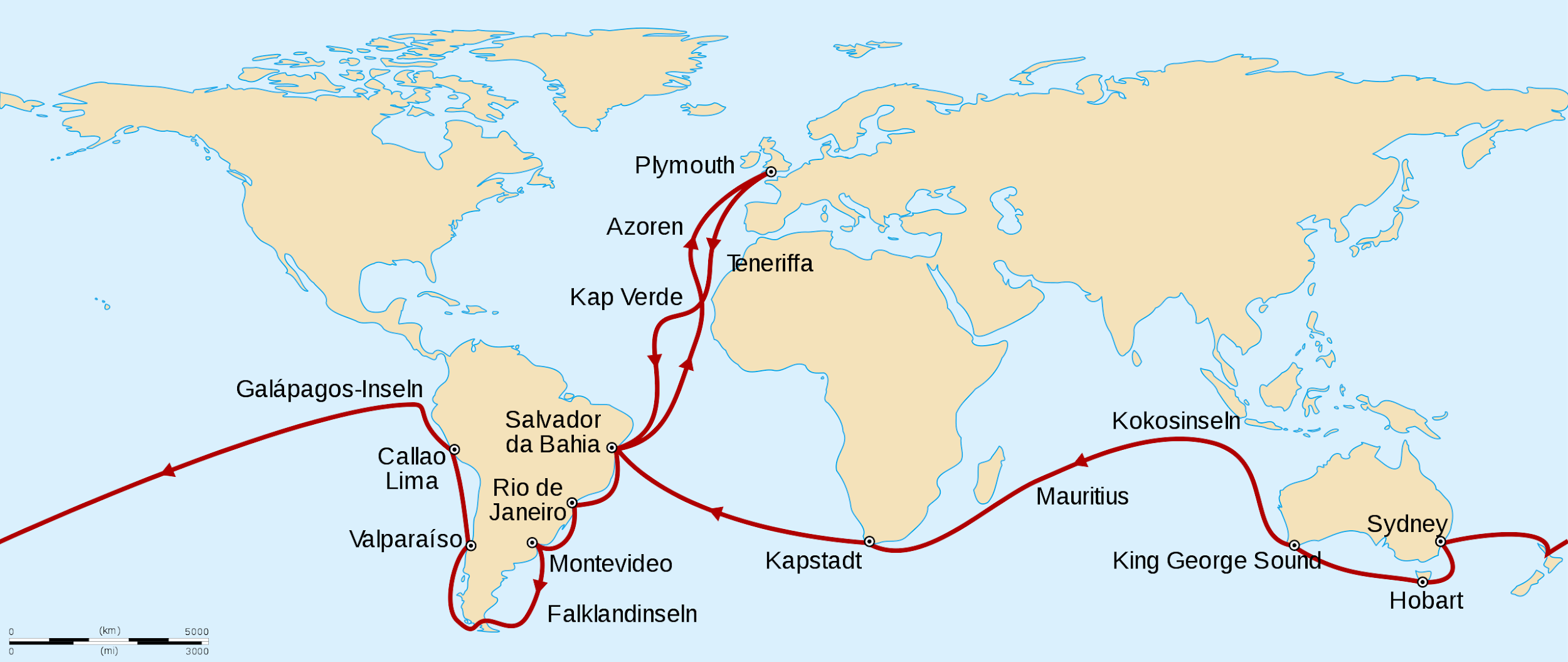
Discussing all of Darwin’s work aboard the Beagle is beyond the scope of this chapter, but his primary interests were in cataloging new varieties of plant and animal life and examining the geology of the places the ship made landfall. Part of Darwin’s success with regard to both ventures was due to his extreme seasickness, which began before the ship even left Plymouth Harbor. It never let up, encouraging Darwin to go ashore at every available opportunity. “In fact, of the nearly five years of the voyage, Darwin was actually on board the ship for just a year and a half altogether” (Costa 2017, 18).
During the voyage, the young Darwin tried to make sense of what he saw through the lens of the scientific paradigms he held when he left England, but he continually made observations that challenged these paradigms. For example, while the Beagle crewmen were charting the coast of Argentina, Darwin conducted fieldwork on land. There he observed species that were new to him, like armadillos. He also collected fossils, including those of extinct armadillos. Meaning, he had found both extant and extinct members of the same species in the same geographic location, which challenged the theory of catastrophism put forth by Cuvier, who argued that each variant of an animal, living or extinct, was its own distinct species (Moore 1993, 144). Darwin also observed geographic variation in the same species all along the east coast of South America, from Brazil to the southern tip of Argentina. He noted that some species were found in multiple localities and differed from place to place. Those living closer to each other exhibited only slight variations, while those living further apart might be cataloged as entirely different species if one did not know better.
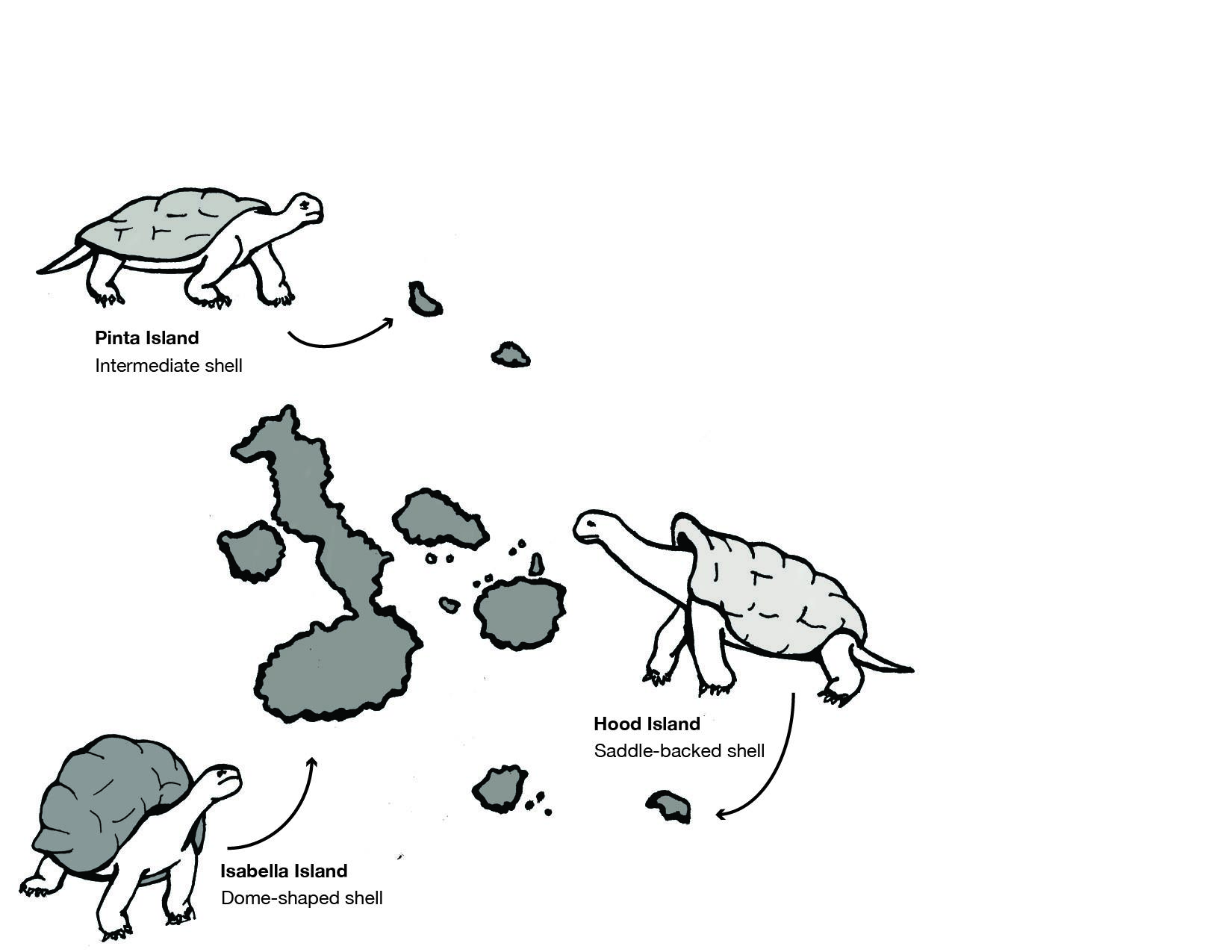
He made similar observations in the Galapagos Islands located off the northwest coast of Ecuador, with regard to giant tortoises and finches (Figure 2.12). A local resident of the islands explained to Darwin that each island had its own variety of tortoise and that locals could discern which island a tortoise came from simply by looking at it. Darwin noted other such examples in both plants and animals, meaning geographic variation was occurring on separate, neighboring islands.
Prevailing views of time argued that variations in living species, and even the fossil armadillos and the living armadillos, were the result of separate creation events. According to this view, each variation, no matter how slight, was a different species. Challenging these ideas would mean challenging not only catastrophism, but the Fixity of Species and other well-accepted ideas of the time. Darwin was aware that he was a young, unestablished naturalist. He was also aware of the ruin that befell Lamarck when his theories were rejected. Lastly, Lyell, who was a good friend of Darwin’s, rejected evolution altogether. It is no wonder that Darwin published a great deal about the geological and fossil data he collected when he returned from the voyage, but not his early hypotheses about evolution.
Upon Darwin’s return to England, it took another twenty years of data collection and experimentation before he was ready to share his conclusions about evolution. Much of this work was conducted at Down House, his home of forty years, where he performed all sorts of experiments that laid the groundwork for his ideas about evolution. Darwin’s home was his laboratory, and he engaged the help of his children, neighbors, friends, and servants in collecting, dissecting, and experimenting. At one point in the 1850s, sheets of moistened paper covered with frogs eggs lined the hallways of the house, while flocks of sixteen different pigeon breeds cooed outside, glass jars filled with salt water and floating seeds filled the cellar, and the smell of dissected pigeon skeletons pervaded the air inside the house. There were also ongoing experiments in the yard, including piles of dissected flowers, beekeeping, and fenced-off plots of land where seedlings were under study. Darwin was a keen experimental scientist, observer, and a prolific writer and presenter of scientific papers. He regarded his work as “one long argument” that never really ended. In fact, Darwin published ten books after On the Origin of Species, addressing such far-ranging topics as animal behavior, orchids, and domestication, among others (Costa 2017).
Darwin may not have published Origins in 1859 had it not been for receiving a paper in June of 1858 from Alfred Russel Wallace, an English naturalist working in Malaysia, espousing the same ideas. Wallace had sent the paper to Darwin asking if it was worthy of publication and requesting he forward it to Lyell and the English botanist, Joseph Hooker. Darwin wrote to Lyell and Hooker about Wallace’s paper, entitled On the Tendency of Varieties to Depart Indefinitely from the Original Type. In recognition that both Wallace and Darwin had arrived at the same discovery, a “joint” paper composed of four parts (none of them actually coauthored) was read to the Linnaean Society by Lyell, then secretary of the Society, on July 1, 1858, and published on August 20. Darwin published On the Origin of Species 15 months later. (The original composite paper read before the Linnaean society is available to read for free from the Alfred Russell Wallace Website, on the 1858 Darwin-Wallace paper page.)
The Mechanism of Natural Selection
Let us take a moment here to explore the mechanism of natural selection in more detail. Before we begin, it is important to recognize that Darwin defined evolution as descent with modification, by which he meant that species share a common ancestor yet change over time, giving rise to new species. Descent with modification refers to the fact that offspring from two parents look different from each of their parents, and from each other, meaning they descend with slight differences (“modifications”). If you have ever observed a litter of puppies or a field of flowers and stopped to examine each individual closely, you have seen that each differs from the next, and none look exactly like their parents. These variations are random, not specific, and may or may not be present in the following generations.
Darwin struggled to explain why some slight differences were preserved over time, while others were not. He turned to what he knew of animal breeding (artificial selection) for an explanation (Richards 1998). Darwin bred different breeds of pigeons at Down House, carefully documenting phenotypic differences across generations, including slight anatomical variations he observed through dissection. He also grew and crossbred species of flowers and dissected those too. Darwin was also very fond of hunting and of hunting dogs. In an early draft of his theory on speciation, he used greyhounds as an example of adaptation and selection, “noting how its every bone and muscle, instinct and habit, were fitted to run down hare (rabbits) (University of Cambridge n.d.).” In each case of plant and animal breeding Darwin observed, he noted that humans were selecting variants in each generation that had characteristics humans desired (i.e., sweetness of fruits, colors of flowers, fur type and color of animals). Breeders then continually bred plants and animals with the desired variants, over and over again. These small changes added up over time to create new species of plants and breeds of animals. Darwin also noted that artificial selection does not necessarily render plants or animals better adapted to their original environments. The characteristics humans desire often result in plants less likely to survive in the wild and animals more likely to suffer from certain behavioral or health problems. One has only to examine high rates of hip dysplasia in several modern breeds of dogs to observe what Darwin was referring to.
From his studies of artificial selection, Darwin drew the conclusion that nature also acts upon variations among members of the same species. Instead of human intervention, the forces of nature, such as heat, cold, predation, disease, and now climate change, determine which offspring, with which variants, survive and reproduce. These individuals then pass down these favorable variants to their own offspring. In this way, nature selects for traits that are beneficial within a particular environment and selects against traits that are disadvantageous within a particular environment. Over many generations, populations of a species become more and more adapted (or, in evolutionary terms, “fit”) for their specific environments. Darwin named this process natural selection.
This theory explained the variations in tortoises Darwin had observed years earlier in the Galapagos Islands (see Figure 2.12). Tortoises who lived on larger islands with lush vegetation to feed on were larger than those on smaller islands. They also had shorter necks and dome-shaped shells as their food was close to the ground. Tortoises on smaller, drier islands fed on cacti, which grew much taller. These tortoises had longer necks, longer front legs, and saddle-shaped shells, which allowed them to successfully stretch to reach the edible cactus pads that grew on the tops of the plants. How did these observable differences in the two tortoise populations emerge? Darwin would argue that, over time, small, random variations in the tortoises were differentially selected for by the distinct natural environments on different islands.
In addition to the biogeographical evidence Darwin offered from his research aboard the Beagle, as well as the evidence he documented from the artificial selection of plants and animals, he also relied, where possible, on fossil evidence. One example, mentioned above, were the fossil findings of extinct armadillos in Argentina in the same locations as living armadillos. Unfortunately, as Darwin himself noted, the geological record was incomplete, most often missing the transitional fossil forms that bridge extinct and living species. That issue has since been resolved with scientific research in geochronology and paleontology, among other fields. It is now well-established that life is far more ancient than was believed in Darwin’s time and that these ancient forms of life were the ancestors to all life on this planet (Kutschera and Niklas 2004).
What Darwin was Missing
Although the theory of evolution by natural selection gained traction in scientific circles in the decades after Darwin’s publication of Origins, he was never able to discover the mechanisms that caused variation within members of the same species or the means by which traits were inherited. This began later in 1892 with the publication of The Germ-Plasm: A Theory of Heredity by August Weismann, the same Weismann of the mouse tail experiment presented earlier in this chapter. In his book, Weissman proposed a theory of germ-plasm, which was a precursor to the later discovery and understanding of DNA. Weismann specialized in cytology, a branch of biology devoted to understanding the function of plant and animal cells. Germ-plasm, he proposed, was a substance in the germ cells (what we would call gametes, or sex cells, today) that carried hereditary information. He predicted that an offspring inherits half of its germ-plasm from each of its parents, and claimed that other cells (e.g. somatic, or body, cells) could not transmit genetic information from parents to offspring. This thereby erased the possibility that acquired traits (which he argued resided in somatic cells) could be inherited (Zou 2015). This contribution to evolutionary theory was an important step toward understanding genetic inheritance, but a connection between genetics and evolution was still lacking.
A series of lectures by a deceased Augustinian monk named Gregor Mendel (1822–1884), originally published in 1865, changed that perspective (Moore 1993, 285). Although Darwin was unknown to Mendel, he began a series of experiments with pea plants shortly after the publication of Darwin’s Origins, aiming to add to evolutionary understandings of heredity. As Mendel bred different generations of pea plants that had differences in seed shape and color, pod shape and color, flower position, and stem length, he documented consistent expression of some variations over others in subsequent generations. He meticulously documented the statistics of each crossing of plants and the percentages of phenotypes that resulted, eventually discovering the concept of dominance and recessiveness of characteristics, as will be seen in chapter 3. The recognition of the importance of Mendel’s work began with its rediscovery by Hugo de Vries and Carl Correns, both of whom were working on hypotheses regarding heredity in plants and had arrived at conclusions similar to Mendel’s. Both published papers supporting Mendel’s conclusions in 1900 (Moore 1993). Research into the inheritance of characteristics continued through the next three decades, and by the close of the 1930s, no major scientific questions remained regarding the transmission of heredity through genes, although what genes did and what chemicals they were made of were still under investigation.
The Modern Synthesis refers to the merging of Mendelian genetics with Darwinian evolution that took place between 1930 and 1950. The basic principles of the synthetic theory were influenced by scientists working in many different fields, including genetics, zoology, biology, paleontology, botany, and statistics. Although there were differences of opinion among them, evolution came to be defined as changes in allele frequencies within populations. Genetic mutations, changes in the genetic code that are the original source of variation in every living thing, were believed to be random, the sources of phenotypic variation, and transmitted through sexual reproduction. These assertions were supported by a growing body of field and laboratory research, as well as new work in mathematics in the field of population genetics that defined evolution as numerical changes in gene frequencies within an interbreeding population from one generation to the next (Corning 2020). These changes in gene frequencies were argued to be the result of natural selection, mutation, migration (gene flow), and genetic drift, or random chance. Empirical research and mathematics demonstrated that very small selective forces acting over a relatively long time were able to generate substantial evolutionary change, including speciation (Plutynski 2009). Thus, the Modern Synthesis encompassed both microevolution, which refers to changes in gene frequencies between generations within a population, and macroevolution, longer-term changes in a population that can eventually result in speciation, wherein individuals from different populations are no longer able to successfully interbreed and produce viable offspring.

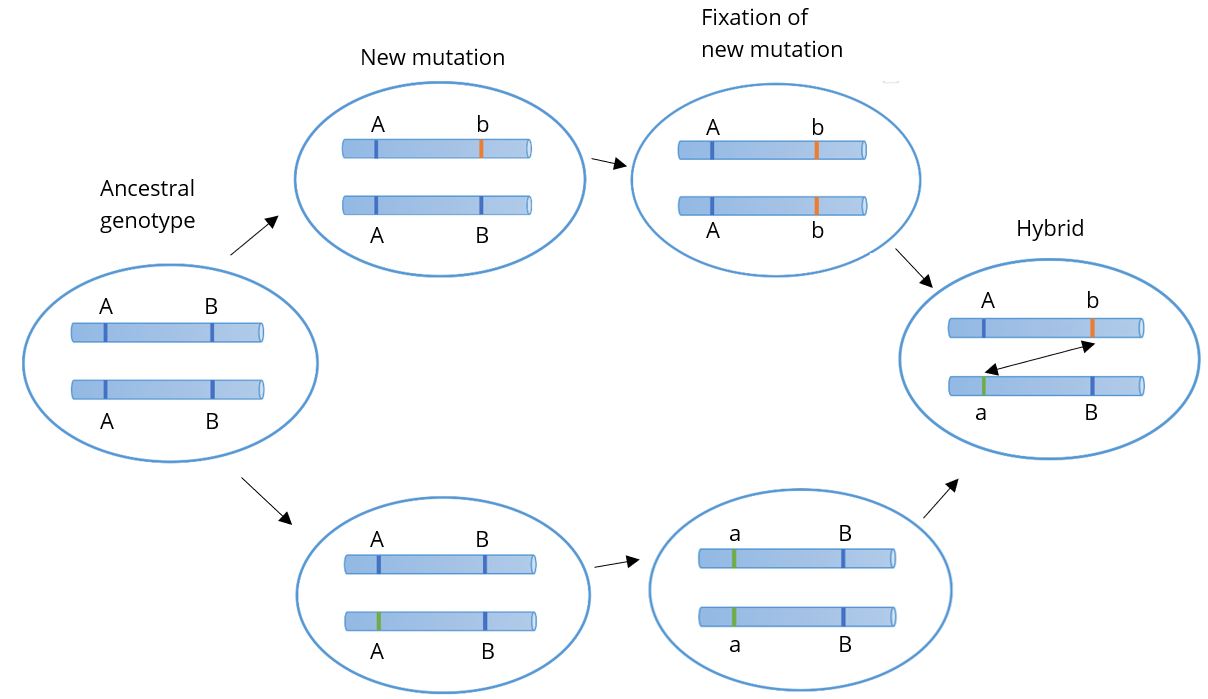
Genetics and the Origin of Species, published in 1937 by Theodosius Dobzhansky (Figure 2.13), was a cornerstone of the modern synthesis, applying genetics to the study of natural selection in wild populations, appealing to both geneticists and field biologists. Dobzhansky was interested in speciation, particularly in finding out what kept one species distinct from another and how speciation occurred. His research involved fruit flies, the species Drosophila pseudoobscura. At the time he began in the 1920s, biologists assumed all members of the same species had nearly identical genes. Dobzhansky traveled from Canada to Mexico capturing wild members of D.pseudoobscura, discovering that different populations had different combinations of alleles (forms of a gene) that distinguished them from other populations, even though they were all members of the same species. What, then, led to the creation of new species? Dobzhansky realized it was sexual selection. Members of the same species are more likely to live among their own kind and to recognize, and prefer, them as mates. Over time, as a result of random mutations, natural selection in a given environment, and genetic drift, meaning random changes in allele frequencies, members of the same population accumulate mutations distinct to their own gene pool, eventually becoming genetically distinct from other populations. What this means is that they have become a new species.
From these studies, Dobzhansky and others developed the Bateson-Dobzhansky-Muller model, also known as Dobzhansky-Muller model (Figure 2.14). It is a model of the evolution of genetic incompatibility. Combining genetics with natural selection, the model is important in understanding the role of reproductive isolation during speciation and the role of natural selection in bringing it about. Due to sexual selection (mate preference), populations can become reproductively isolated from one another. Eventually, novel mutations may arise and be selected for in one or both populations, rendering members of each genetically incompatible with the other, resulting in two distinct species.
Special Topic: Evolution and Natural Selection Observable Today

Although this chapter primarily focuses on the past, it is important to remember that natural selection and evolution are still ongoing processes. Climate change, deforestation, urbanization, and other human impacts on the planet are influencing evolution among many contemporary species of plants and animals. One such example occurs among crested anoles (Anolis cristatellus), small lizards of the Caribbean jungle that are increasingly making their home in cities (Figure 2.15).
As urban sprawl continues across the planet, shrinking the availability of wilderness habitat, many wild species have come to make their homes in cities. “Urbanization has dramatically transformed landscapes around the world—changing how animals interact with nature, creating “heat islands” with higher temperatures, and hurting local biodiversity. Yet many organisms survive and even thrive in these urban environments, taking advantage of new habitats created by humans (National Science Foundation 2023). A recent example of lizards in Puerto Rico demonstrates evolution happening quickly in both behavior and genes that has come about as a result of the pressures of urban life (Winchell et al. 2023). Crested anoles, who once lived only in forests, now scurry around towns and cities throughout the Caribbean. As a result of having to sprint across large open spaces, like hot streets and parking lots, they have developed longer limbs. City-living lizards also now sport longer toe pads with special scales that allow them to cling to smooth surfaces, like windows and walls (as well as the plastic patio furniture pictured in Figure 2.15), rather than to the rough surfaces of bark and rock that their forest-living relatives climb. These adaptations enhance their ability to escape predators and survive in cities.
Researchers were curious to see if these changes were the result of genetic changes in urban populations, so they captured 96 male lizards in three Puerto Rican regions and compared their genomes to each other and to forest specimens in each location. They found that members of the three city-living populations were genetically distinct from each other, as well as from forest populations in their respective regions. In total, 33 genes in the urban lizards’ genomes were different from their forest-living counterparts and were linked to urbanization. These changes are estimated to have occurred just within the last 30 to 80 generations, suggesting that selective pressures related to survival in urban environments is strong. As study coauthor Kristen Winchell put it, “We are watching evolution as it is unfolding” (National Public Radio 2023). (If you are interested in hearing more about the study, see “How Lizards Adapt to Urban Living,” an episode of Science Sessions, a free podcast from the Proceedings of the National Academy of Sciences (PNAS 2023) featuring Dr. Winchell and her work.)
Misconceptions About Evolution Through Natural Selection
After many years of teaching about evolution and natural selection, it continues to surprise me how many misconceptions exist about how the process works. If you do a web image search for “human evolution,” the following image is likely to appear (Figure 2.16).

What is wrong with this picture? First, it implies that humans evolved from chimpanzees, which is incorrect. Although, as primates, we share a common ancestor very far back in time, we split from other primates, including our closest relatives, the nonhuman apes, several million years ago. This image also suggests that evolution is gradual and progressive; that it is intentional and directional; and that there is an end to it—a stopping point. As you will be learning, evolution takes place in fits and starts, depending on the physical environment, changes in climate, food supply, predation, reproductive success, and other factors. It is also not intentional, in the sense that there is no predetermined end; in fact, if environmental conditions change, species can evolve in different directions or even go extinct. Evolution also does not necessarily progress in the same direction over time. One example is the eel-like creature Qikiqtania wakei that lived 375 million years ago. It was originally a fish that evolved to walk on land, then evolved to live back in the water. Early tetrapods like Qikiqtania were likely spending more and more time out of the water during this period. The arrangement of bones and joints in their fins was starting to resemble arms and legs, which would have allowed them to prop themselves up in shallow water and survive on mudflats. Qikiqtania’s skeletal morphology, however, suggests that it then evolved from having rudimentary fingers and toes back to fins that allowed them to again swim in open water (Stewart et al. 2022).
There is also the misperception that natural selection can create entirely new anatomical structures out of thin air in response to changes in environmental pressures. For example, when asked if they can think of ways in which modern humans are continuing to evolve biologically, students often postulate that, as a result of climate change, humans might rapidly develop gills, webbed hands and feet, and learn to breathe underwater in response to rising sea levels. Unfortunately, natural selection can only act on slight variations in anatomy that are already present, and we have no rudimentary physiological system for breathing underwater. Given that natural selection can only act upon existing variation, humans have evolved in such a way that many parts of our bodies are prone to injury. Our knees are one example. The anterior cruciate ligament (ACL) in our knees is “vulnerable to tearing in humans because our upright bipedal posture forces it to endure much more strain than it is designed to” (Lents 2018, 23). When our ancestors made the transition from quadrupedalism to upright walking, we shifted from four bent legs to two straight legs, relying more on our bones than our muscles to support our weight. This is functional for normal walking and running in a straight line, but sudden shifts in direction and momentum, combined with the sizes and weights of modern humans, result in tears in an ACL that is simply not strong enough to bear the stress. If evolution had the capability to engineer a knee from scratch, it would look quite different, and any ligaments involved would likely be larger, stronger, and more flexible. For an interesting look at what anatomically modern humans might look like if we had evolved to withstand the stresses our bodies undergo in our present environment, see “This is what the perfect body looks like – according to science,” which was proposed by biological anthropologist Alice Roberts (Harrison 2018).
Another misperception about evolution is that some species are “more evolved” than others. Every species currently alive on the planet today is the result of millennia of natural selection that has rendered current members of that species well-adapted to their respective environments. Humans are no more “evolved” than fruit flies or yeast. What sets us apart are our cultural and technological abilities, which have allowed us to successfully survive in a wide variety of physical environments, many of which are now becoming too hot, too wet, or too dry to sustain human life without a great deal of technological intervention (IPCC 2022).

There is also some confusion about what “fitness” actually means and a failure to grasp that it changes as environmental conditions change. Evolutionary “fitness” is different from physical fitness. “Fitness” in evolutionary terms refers to an individual’s ability to survive and reproduce viable offspring who also survive and reproduce. Evolutionary fitness and reproductive success are highly dependent on specific environmental conditions, which can shift over time, greatly affecting the relative fitness of individuals in a population. Recent research on the impacts of climate change on dragonflies will serve to illustrate the point (Figure 2.17).
Pictured here is a male dragonfly, who, you will notice, has distinctive black markings on its wings. This is due to melanization. Males control breeding, and those with more ornamentation tend to attract more mates and to successfully ward off male competitors. Higher levels of melanization, however, have negative consequences for males in warming climates. The black markings absorb heat, elevating body temperatures, which can cause overheating, reduce male fighting ability, and even lead to death (Moore et al. 2021). Females are not as adversely affected because they spend more time in shaded areas, while males are more often flying in sunlit areas, fending off rivals. However, as highly melanized males become less viable, wing coloration is undergoing selection in males. In other words, what constitutes being “fit” for males has changed, favoring those who have fewer of the black markings and, therefore, are less negatively impacted by warming temperatures. Note that natural selection acts on individuals, “selecting” those who happen to be fit for particular environmental conditions at a particular point in time. Evolution, though, happens at the level of the population. If the climate continues to warm, populations of dragonflies who inhabit warming areas will increasingly exhibit less ornamentation in males.
Lastly, natural selection can only act on characteristics that influence reproductive success. Deleterious traits that have nothing to do with one’s ability to reproduce and successfully rear offspring to reproductive age will continue to be passed on. For example, the author of this chapter is a natural redhead, and redheads are predisposed genetically to a number of conditions that can negatively affect health (Colliss Harvey 2015), but some of these conditions are not diagnosed until later in life. One example is Parkinson’s disease (Chen et al. 2017), which is a degenerative neurological disorder. The average age of diagnosis of Parkinson’s is 60 years of age, meaning redheads may encounter such a diagnosis well past childbearing age, having already passed on the genetic predisposition. Thus, Parkinson’s disease cannot be selected out from the redhead family tree.
Are We Still Evolving?
After reading this chapter, many students are curious to know if humans are still evolving. The answer is yes. As a species, we continue to respond to selective pressures biologically and culturally. This final section will focus on three contemporary examples of human evolution. Before beginning, let’s review the conditions necessary for natural selection to operate on a trait. First, the trait must be heritable, meaning it is transmitted genetically from generation to generation. There must also be variation of the trait within the population and the trait must influence reproductive success. Three examples of traits that meet these criteria are immunity to the Human Immunodeficiency Virus (HIV), height, and wisdom teeth (Andrews, Kalinowski, and Leonard 2011).
AIDS is a potentially fatal infectious disease caused by HIV, a zoonosis believed to be derived from Simian Immunodeficiency Viruses (SIVs) found in chimpanzees and monkeys and most likely transmitted to humans through the butchering of infected animals (Sharp and Hahn 2011). In total, 40 million people have died from AIDS-related illnesses since the start of the global epidemic in the 1980s. There were 38.4 million people around the world living with AIDS as of 2021, including 1.5 million new cases and 650,000 deaths in that year alone (UNAIDS 2021). A disease causing this level of morbidity and mortality represents a major selective pressure, especially given that infection can occur before birth (Goulder et al. 2016), thereby affecting future reproductive success.
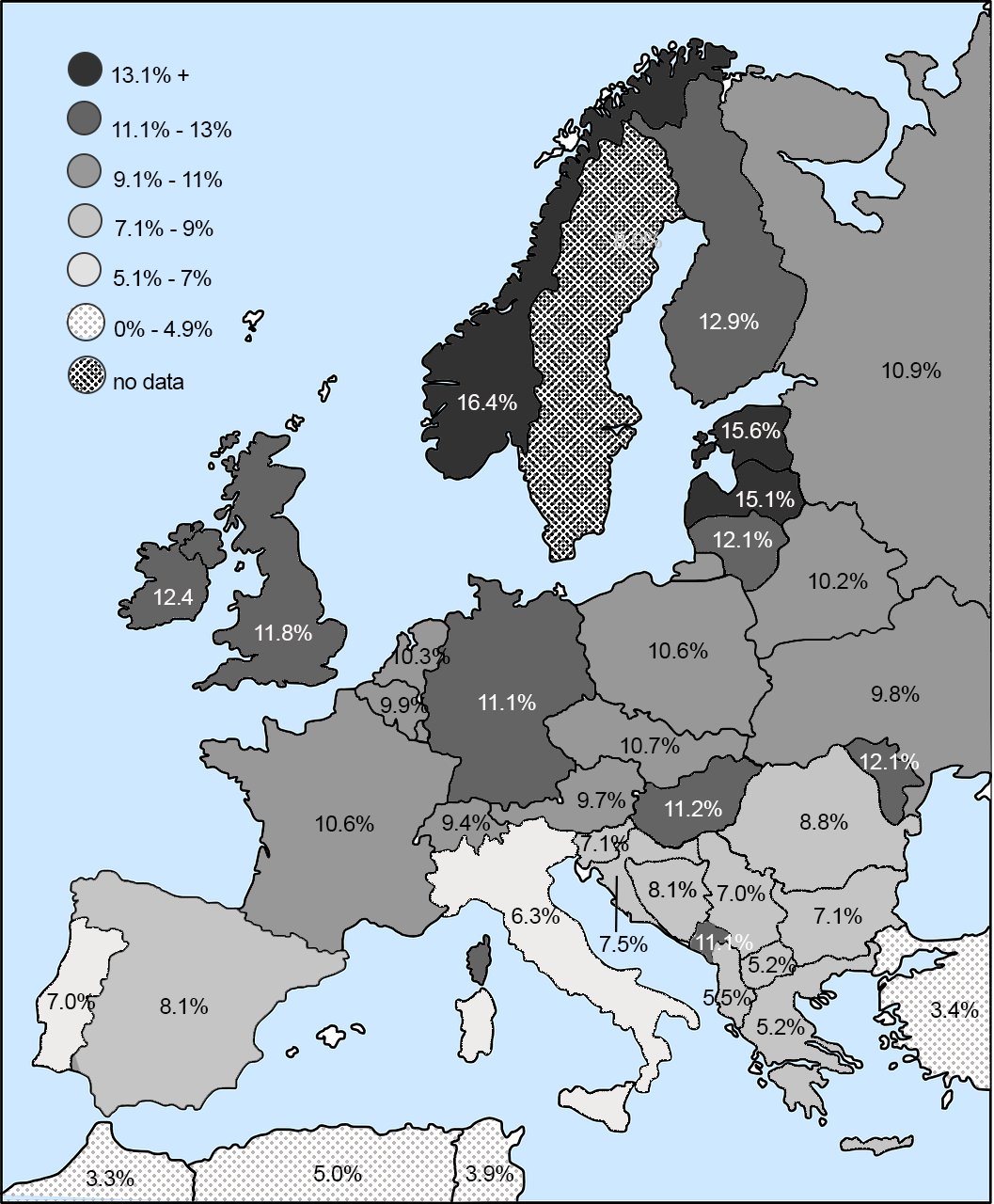
The majority of people in the world are highly susceptible to HIV infection, but some are not. These latter individuals are homozygous for a rare, recessive allele at the CCR5 locus that makes them immune to HIV. Heterozygotes who inherit a single copy of this allele are more resistant to infection and, when infected, the disease takes longer to progress in the event that they are infected. The mechanism by which the allele prevents infection involves a 32-base pair deletion in the DNA sequence of the CCR5 gene, creating a nonfunctioning receptor on the surface of the cell that prevents HIV from infecting the cell. The allele is inherited as a simple Mendelian trait, and there is variation in its prevalence, ranging as high as 14% of the population in northern Europe and Russia (Novembre, Galvani, and Slatkin 2005; see Figure 2.19). What is interesting about the allele’s geographic distribution is that it does not map onto parts of the world with the highest rates of HIV infection (Figure 2.20), suggesting that AIDS was not the original selective pressure favoring this allele (see Figures 2.19 and 2.20).
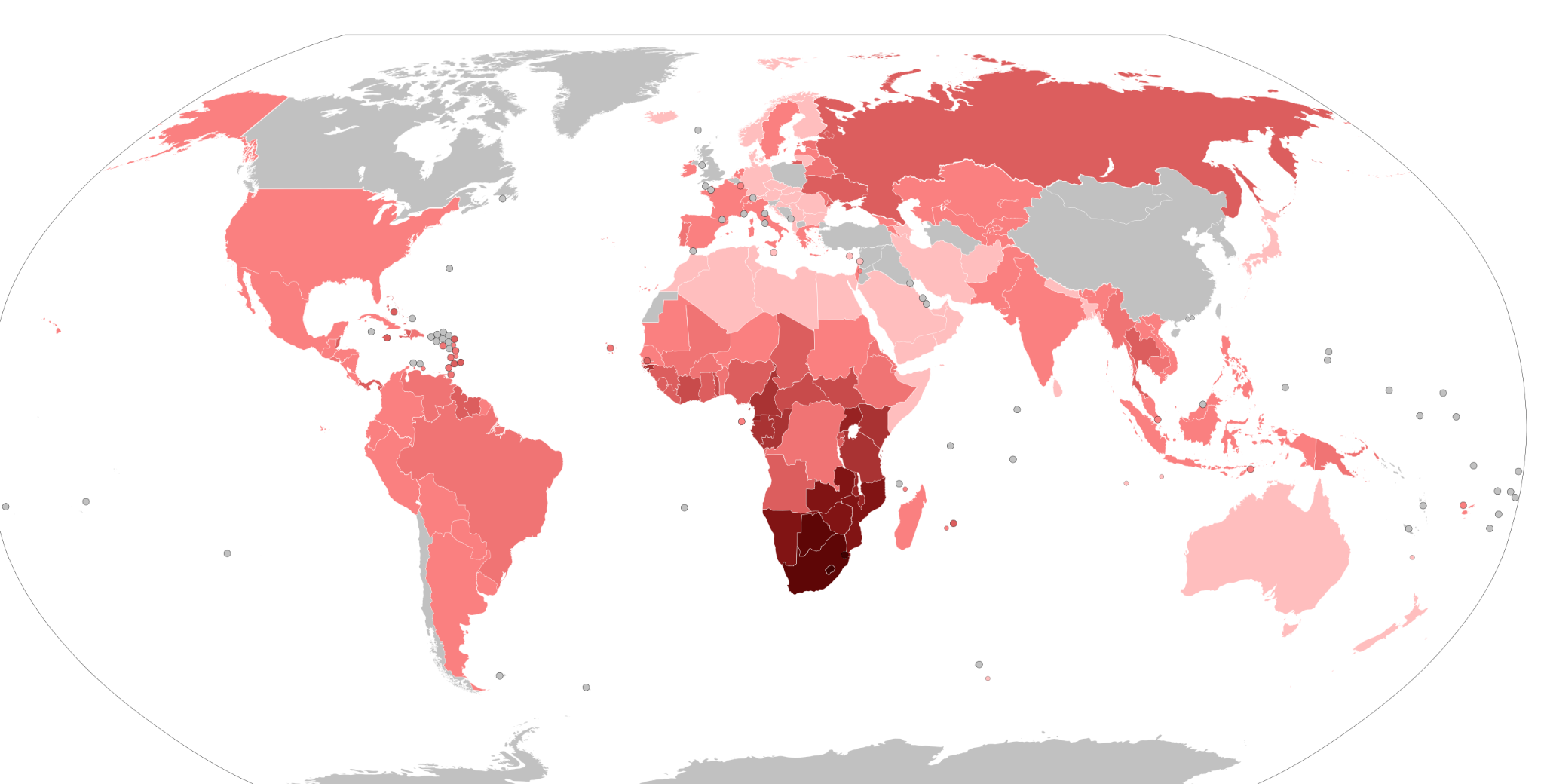
Given its current geographic distribution, the bubonic plague, which ravaged Europe repeatedly from the 14th to the 19th centuries (Pamuk 2007), was initially proposed as the selective agent. Subsequent research suggests smallpox, which killed up to 400,000 people annually in 18th-century Europe (Hays 2005), was more likely the selective pressure (Novembre, Galvani, and Slatkin 2005). Given the mortality rates for smallpox (Crosby 2003), an allele that conferred immunity was highly advantageous, as it is for those faced with the threat of HIV infection today.
Height is another example of a trait experiencing selective pressure. If you have ever toured a historical site, you have likely hit your head on a doorframe or become claustrophobic trying to squeeze down a narrow hallway under a lower-than-average ceiling. It is not your imagination. Humans have gotten taller in recent centuries. In fact, the average height of people in industrialized nations has increased approximately 10 centimeters (about four inches) in the past 150 years. This increase has been attributed to improvements in nutrition, sanitation, and access to medical care (Hatton 2014). But this is only part of the story.
Height is highly heritable. Studies indicate 80% of variation in height within populations is due to genetics, with 697 different genetic variances identified as having an effect on adult stature (Devuyst 2014). Multiple studies also demonstrate a positive relationship between height and reproductive success for men (Andrews, Kalinowsky, and Leonard 2011). This is primarily due to sexual selection and nonrandom mating, namely women’s preferences for taller men, which may explain why height is one characteristic men often lie about on dating websites (Guadagno, Okdie, and Kruse 2012). Sexual selection also plays out with regard to economic success in Western cultures, with taller men more likely to be in higher-level positions that pay well. Research demonstrates an inch of height is worth an additional $789 per year in salary, meaning a man who is six feet tall will earn on average $5,525 more per year than an identical man who is five foot five purely due to heightism bias (Gladwell 2007). Over the course of a career, this can add up to hundreds of thousands of dollars, likely allowing taller men to attract more potential mates, increasing their reproductive success.
Wisdom teeth are also undergoing evolutionary pressure. Have you or anyone in your family had their wisdom teeth removed? While it can be a painful and expensive process, it is a common experience in Western nations. It begs the question as to why there is no longer room in our mouths for all of our teeth? Biological anthropologist Daniel Lieberman offers several reasons, including that modern humans are growing faster and maturing earlier, which could be leading to impaction if skeletal growth takes place faster than dental growth. He also argues that the soft diets many modern humans consume generate insufficient strain to stimulate enough growth in our jaws to accommodate all of our teeth. Lastly, as the human brain has expanded over the past hundreds of thousands of years, it is taking up more space in the skull, causing the jaw to shrink, leaving no room for third molars (Lieberman 2011).
Conversely, do you know anyone whose wisdom teeth never came in? That is fairly common in some populations, suggesting evolutionary pressure favoring the absence of wisdom teeth has been culturally influenced. The oldest fossil evidence of skulls missing third molars was found in China and is 300,000 to 400,000 years old, suggesting the earliest mutation selecting against the eruption of wisdom teeth arose in Asia (Main 2013). Since that time, jaws have continued to decrease in size to the point they often cannot accommodate third molars, which can become impacted, painful, and even infected, a condition physical anthropologist Alan Main argues might have interfered with the ability to survive and reproduce in ancestral populations (Main 2013). As we have learned, a mutation that positively influences reproductive success—such as being born without the trait to develop wisdom teeth—would likely be selected for over time. Evidence in modern humans suggests that this is the case, with 40% of modern Asians and 45% of Native Alaskans and Greenlanders (populations descended from Asian populations) lacking wisdom teeth. The percentage among those of European descent ranges from 10 to 25% and for African Americans is 11% (Main 2013). Later chapters of this textbook emphasize that directional selection progresses along a particular path until the environment changes and a trait is no longer advantageous. In the case of wisdom teeth, the ability of modern dentistry to preempt impaction through surgery may, in fact, be what is preventing natural selection from doing away with wisdom teeth altogether.
Key Developments in Evolutionary Thought
|
4th century BCE |
Aristotle (384–322 BCE) |
“Founder of Biology.” Publishes History of Animals, a biological classification system of over 500 animals based on structure, physiology, reproduction, and behavior. Also creates the “Great Chain of Being,” ranking species and placing humans closest to God. |
|
8th–9th century CE |
Al-Jahiz (776–868 CE) |
Writes seven-volume Book of Animals, which includes animal classifications and food chains. Introduces concept of biological evolution and its mechanisms. |
|
1011–1021 |
Ibn al-Haythem (965–1040 CE) |
“Father of Modern Optics.” Uses experimental science to catalog how vision works and discovers laws of reflection and refraction. Publishes Book of Optics and invents camera obscura, the foundation for modern photography. |
|
1620 |
Francis Bacon (1561–1626) |
“Father of Empiricism.” Publishes The Novum Annum, formulating the scientific method based on observation and inductive reasoning. |
|
1686 |
John Ray (1627–1705) |
First to publish a biological definition of species in History of Plants. |
|
1749 |
Comte de Buffon (1707–1788) |
Publishes Histoire Naturelle, comparing anatomical structures across species using methods still in use today. Inspires Lamarck and Cuvier. |
|
1758 |
Carl von Linne (Carolus Linnaeus) (1707–1778) |
Introduces system of binomial nomenclature. Publishes Systema Naturae, the tenth edition of which introduces the designation Homo sapiens for humans. |
|
1788 |
James Hutton (1726–1797) |
“Father of Geology.” Publishes Theory of the Earth; introduces idea of Deep Time; explains how features of the earth were formed through the actions of rain, wind, rivers, and volcanic eruptions. |
|
1798 |
Thomas Malthus (1766–1834) |
Economist and “Father of Statistics.” Publishes An Essay on Population; introduces concept of carrying capacity; explains how populations outstrip the food supply, leaving some individuals to die off; inspires Darwin’s idea of “natural selection.” |
|
1809 |
Jean-Baptiste Lamarck (1744–1829) |
Publishes theory of the Inheritance of acquired characteristics; is the first Western scientist to propose a mechanism explaining how traits change in species over time and to recognize the importance of the physical environment in acting on species and their survival. |
|
1810 |
Georges Cuvier (1769–1832) |
Paleontologist/comparative anatomist; proved species went extinct; proposed the Theory of Catastrophism. |
|
1830 |
Charles Lyell (1797–1875) |
Establishes geology as a science. Publishes first edition of The Principles of Geology (1830–33); issuing 12 total editions in his lifetime, each updated according to new scientific data. |
|
1858 |
Alfred Russel Wallace (1823–1913) |
Sends scientific paper to Darwin titled “On the Tendency of Varieties to Depart Indefinitely from the Original Type,” essentially espousing the concept of natural selection; a reading of the papers by both Wallace and Darwin to the Linnaean Society is conducted by Lyell. |
|
1859 |
Charles Darwin (1809–1882) |
Publishes On the Origin of Species by Means of Natural Selection (1859). |
|
1865 |
Gregor Mendel (1822–1884) |
Publishes Experiments in Plant Hybridization (1865), outlining the fundamentals of genetic inheritance. |
|
1889 |
August Weismann (1834–1914) |
Publishes Essays Upon Heredity (1889), disproving the inheritance of acquired characteristics. Publishes The Germ Plasm (1892), postulating an early idea of inheritance through sexual reproduction. |
|
1937 |
Theodosius Dobzhansky (1900–1975) |
One of the founders of the Modern Synthesis of biology and genetics. Publishes Genetics and the Origin of Species (1937). Documents a genetic model of speciation through reproductive isolation. |
Summary
Firstly, it is important to recognize that the global discourse on evolutionary thought emerged from a Western European colonial legacy; often centering eurocentric perspectives while overlooking other intellectual traditions. This legacy has resulted in a lasting influence in how the knowledge was–and continues to be–structured and understood.
From its earliest ideas to today’s genomic research, evolutionary thought has demonstrated that species, including our own, are not static, but are part of ongoing processes shaped by variation, natural selection, and shifting conditions. A persistent misconception is that evolution implies progress toward more advanced or ‘better’ forms; in reality, it reflects context-specific adaptations that enhance survival and reproduction. Humans exemplify this ongoing process, adapting through traits such as resistance to disease, tolerance of new foods, and responses to rapidly changing modern environments.
Review Questions
- Summarize the major scientific developments that led to the formulation of the theory of natural selection.
- Explain how natural selection operates and how it leads to evolution in populations.
- Explain the importance of genetics to an understanding of human evolution.
- Have you observed current examples of evolution taking place where you live? In which species? Which forces of evolution are involved?
Key Terms
Allele: A nonidentical DNA sequence found in the same gene location on a homologous chromosome, or gene copy, that codes for the same trait but produces a different phenotype.
Artificial selection: The identification by humans of desirable traits in plants and animals, and the subsequent steps taken to enhance and perpetuate those traits in future generations.
Binomial nomenclature: A system of classification in which a species of animal or plant receives a name consisting of two terms: the first identifies the genus to which it belongs, and the second identifies the species.
Carrying capacity: The number of living organisms, including animals, crops, and humans, that a geographic area can support without environmental degradation.
Catastrophism: The theory that the Earth’s geology has largely been shaped by sudden, short-lived, violent events, possibly worldwide in scope. Compare to uniformitarianism.
Comparative anatomy: Georges-Louis Leclerc’s technique of comparing similar anatomical structures across different species.
Creationism: The belief that the universe and all living organisms originate from specific acts of divine creation, as in the Biblical account, rather than by natural processes such as evolution.
Empiricism: The idea that all learning and knowledge derives from experience and observation. It became prominent in the 17th and 18th centuries in western Europe due to the rise of experimental science.
Evolution: In a biological sense, this term refers to cumulative inherited change in a population of organisms through time. More specifically, evolution is defined as a change in allele (gene) frequencies from one generation to the next among members of an interbreeding population.
Extant: Still in existence; surviving.
Extinct: Said of a species, family, or other group of animals or plants that has no living members; no longer in existence.
Fixity of Species: The idea that a species, once created, remains unchanged over time.
Gene: A sequence of DNA that provides coding information for the construction of proteins.
Genetic drift: Random changes in allele frequencies within a population from one generation to the next.
Gene flow: The introduction of new genetic material into a population through interbreeding between two distinct populations.
Gene pool: The entire collection of genetic material in a breeding community that can be passed from one generation to the next.
Genotype: The genotype of an organism is its complete set of genetic material—its unique sequence of DNA. Genotype also refers to the alleles or variants an individual carries in a particular gene or genetic location.
Hybrid: Offspring of parents that differ in genetically determined traits.
Intelligent design: A pseudoscientific set of beliefs based on the notion that life on earth is so complex that it cannot be explained by the scientific theory of evolution and therefore must have been designed by a supernatural entity.
Macroevolution: Large and often-complex changes in biological populations, such as species formation.
Microevolution: Changes in the frequency of a gene or allele in an interbreeding population.
Modern synthesis: The mid–20th century merging of Mendelian genetics with Darwinian evolution that resulted in a unified theory of evolution.
Natural selection: The natural process by which the survival and reproductive success of individuals or groups within an interbreeding population that are best adjusted to their environment leads to the perpetuation of genetic qualities best suited to that particular environment at that point in time.
Phenotype: The detectable or visible expression of an organism’s genotype.
Scientific method: A method of procedure that has characterized natural science since the 17th century, consisting of systematic observation, measurement, experimentation, and the formulation, testing, and modification of hypotheses.
Speciation: The process by which new genetically distinct species evolve from the main population, usually through geographic isolation or other barriers to gene flow.
Species: A group of living organisms consisting of similar individuals capable of exchanging genes or interbreeding. The species is the principal natural taxonomic unit, ranking below a genus and denoted by a Latin binomial (e.g., Homo sapiens).
Uniformitarianism: The theory that changes in the earth’s crust during geologic history have resulted from the action of continuous and uniform processes—such as wind, precipitation, evaporation, condensation, erosion, and volcanic action—that continue to act in the present. Compare to catastrophism.
For Further Exploration
Costa, James T. 2017. Darwin’s Backyard: How Small Experiments Led to a Big Theory. New York: W.W. Norton.
Darwin, Charles. 1905. The Voyage of the Beagle. (Originally published in 1839 as Journal and Remarks). [Author’s note: Several editions exist with different publishers, including illustrated editions, paperback editions, and e-books.]
Moore, John A. 1993. Science as a Way of Knowing: The Foundations of Modern Biology. Cambridge, MA: Harvard University Press.
References
Al-Haytham, Ibn. 1011-1021. Kitāb al-Manāẓir (Book of Optics). Cairo, Egypt.
Al-Jahiz. 776–868 CE. Kitab al-Hayawan (Book of Animals).
Andrews, Tessa M., Steven T. Kalinowski, and Mary J. Leonard. 2011. “Are Humans Evolving? A Classroom Discussion to Change Students’ Misconceptions Regarding Natural Selection.” Evolution: Education and Outreach 4 (3): 456–466.
Aristotle. 384-322 BCE. History of Animals.
Asghar, Anila, Salman Hameed, and Najme Kashani Farahani. 2014. “Evolution in Biology Textbooks: A Comparative Analysis of Five Muslim Countries.” Religion & Education 41 (1). Accessed February 12, 2023. https://www.tandfonline.com/doi/abs/10.1080/15507394.2014.855081.
Associated Press. January 10, 2023. “Forest Lizards Have Genetically Morphed To Survive Life In The City, Researchers Say.” National Public Radio (NPR). Retrieved February 19, 2023 from https://www.npr.org/2023/01/10/1148150056/forest-lizards-genetically-morphed-cities.
Burkhardt, Richard W. 2013. “Lamarck, Evolution, and the Inheritance of Acquired Characters.” Genetics 194 (4): 793–805.
Chen, Xiqun, Danielle Feng, Michael A. Schwartzchild, and Xiang Gao. 2017. “Red Hair, MC1R Variants, and Risk for Parkinson’s Disease—A Meta-Analysis.” Annals of Clinical and Translational Neurology 4 (3): 212–216.https://doi.org/10.1002/acn3.381.
Colliss Harvey, Jacky. 2015. Red: A History of the Redhead. New York: Black Dog & Levanthal.
Corning, Peter A. 2020. “Beyond the Modern Synthesis: A Framework for a More Inclusive Biological Synthesis.” Progress in Biophysics and Molecular Biology 153: 5–12.
Costa, James T. 2017. Darwin’s Backyard: How Small Experiments Led to a Big Theory. New York: W. W. Norton.
Crosby, Alfred W., Jr. 2003. The Columbian Exchange: Biological and Cultural Consequences of 1492. 30th Anniversary Edition. Westport, CT: Praeger.
Darwin, Charles. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. First Edition. London: John Murray.
Darwin, Francis, ed. 2001[1897]. The Life & Letters of Charles Darwin, vol. 2. Honolulu: University Press of the Pacific.
Desilver, Drew. 2017. “U.S. Students’ Academic Achievement Still Lags That of Their Peers in Many Other Countries.” Pew Research Center, February 15. Accessed May 25, 2022. https://www.pewresearch.org/fact-tank/2017/02/15/u-s-students-internationally-math-science/.
Devuyst, Olivier. 2014. “High Time for Human Height.” Peritoneal Dialysis International 34 (7):685–686.
Dobzhansky, Theodosius. 1937. Genetics and the Origin of Species. Columbia University Biological Series (Volume 11). New York: Columbia University Press.
Dunbar-Ortiz, Roxanne. 2014. An Indigenous Peoples’ History of the United States. Boston: Beacon.
El-Zaher, Sumaya. 2018. “The Father of the Theory of Evolution: Al-Jahiz and His Book of Animals.” MVSLIM.com, October 9. Accessed August 27, 2022. https://mvslim.com/the-father-of-the-theory-of-evolution-al-jahiz-and-his-book-of-animals/.
Elshakry, Marwa. 2010. “When Science Became Western: Historiographical Reflections.” ISIS 101 (1). Accessed November 20, 2022. https://www.journals.uchicago.edu/doi/10.1086/652691.
Gladwell, Malcolm. 2007. Blink: The Power of Thinking without Thinking. New York: Back Bay
Guadagno, Rosanna E., Bradley M. Okdie, and Sara A. Kruse. 2012. “Dating Deception: Gender, Online Dating, and Exaggerated Self-Presentation.” Computers in Human Behavior 28 (2): 642–647.
Harmon, Katherine. 2011. “Evolution Abroad: Creationism Evolves in Science Classrooms around the Globe.” Scientific American, March 3. Accessed May 25, 2022. https://www.scientificamerican.com/article/evolution-education-abroad/.
Harrison, Ellie. 2018. “This is What the Perfect Body Looks Like – According to Science.” Radiotimes.com. Accessed June 14, 2023. https://www.radiotimes.com/tv/documentaries/this-is-what-the-perfect-body-looks-like-according-to-science/.
Hatton, Tim. 2014. “Why Did Humans Grow Four Inches in 100 Years? It Wasn’t Just Diet.” The Conversation, May 1. https://theconversation.com/why-did-humans-grow-four-inches-In-100-years-it-wasnt-just-diet-25919.
Hays, J. N. 2005. Epidemics and Pandemics: Their Impacts on Human History. Santa Barbara, CA: ABC-CLIO.
Hutton, James. 1788. Theory of the Earth. Transactions of the Royal Society of Edinburgh, Volume 1. Scotland: Royal Society of Edinburgh.
IPCC. 2022. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, edited by H.-O. Pörtner, D.C. Roberts, M. Tignor, E.S. Poloczanska, K. Mintenbeck, A. Alegría, M. Craig, S. Langsdorf, S. Löschke, V. Möller, A. Okem, and B. Rama. Cambridge: Cambridge University Press. https://www.ipcc.ch/report/ar6/wg2/.
Jones, Stephen. 2017. “Religion, Science, and Evolutionary Theory”[an interview with Salman Hameed]. Podcast, The Religious Studies Project, January 30. Accessed May 29, 2023. https://www.religiousstudiesproject.com/podcast/religion-science-and-evolutionary-theory/.
Kutschera, Ulrich, and Karl J. Niklas. 2004. “The Modern Theory of Biological Evolution: An Expanded Synthesis.” Naturwissenschaften 91: 255–276.
Leclerc, Georges-Louis, Comte de Buffon. 1749-1804. Histoire Naturelle. Volumes 1-36. Paris: Imprimerie Royale (Royal Printing House).
Lents, Nathan H. 2018. Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken Genes. Boston: Houghton Mifflin Harcourt.
Lerner, Lawrence S. 2000. Good Science, Bad Science: Teaching Evolution in the States. Washington, DC: Thomas B. Fordham Foundation.
Lieberman, Daniel E. 2011. The Evolution of the Human Head. Cambridge, MA: Harvard University Press.
Lindberg, David C. 1992. The Beginnings of Western Science: The European Scientific Tradition in Philosophical, Religious, and Institutional Context, 600 B.C. to A.D. 1450. Chicago: The University of Chicago Press.
Linnaeus, Carl. 1736. Systema Naturae. First Edition. Stockholm: Laurentius Salvius.
Love, Shayla. 2020. “A Thousand Years before Darwin, Islamic Scholars Were Writing about Evolution.” VICE World News, October 5. Accessed August 27, 2022. https://www.vice.com/en/article/ep4ykn/a-thousand-years-before-darwin-islamic-scholars-were-writing-about-natural-selection.
Main, Douglas. 2013. “Ancient Mutation Explains Missing Wisdom Teeth.” Live Science, March 13. https://www.livescience.com/27529-missing-wisdom-teeth.html.
Malthus, Thomas Robert. 1798. An Essay on the Principle of Population. London: J. Johnson.
Masci, David. 2019. “For Darwin Day, 6 Facts About the Evolution Debate.” Pew Research Center. Accessed June 14, 2023. https://www.pewresearch.org/short-reads/2019/02/11/darwin-day/.
Masood, Ehsan. 2009. “Islam’s Evolutionary Legacy.” The Guardian, March 1. Accessed February 13, 2023. https://www.theguardian.com/commentisfree/belief/2009/feb/27/islam-religion-evolution-science.
Miller, Jon D., Eugenie C. Scott, Mark S. Ackerman, Belen Laspra, Glenn Branch, Carmelo Polino, and Jordan S. Huffaker. 2022. “Public Acceptance of Evolution in the United States, 1985–2020.” Public Understanding of Science 31 (2): 223–238.
Miller, Jon D., Eugenie C. Scott, and Shinji Okamoto. 2006, August 11. “Public Acceptance of Evolution.” Science 313 (5788): 765-766. DOI: 10.1126/science.1126746.
Moore, John A. 1993. Science as a Way of Knowing: The Foundations of Modern Biology. Cambridge, MA: Harvard University Press.
Moore, Michael P., Kaitlyn Hersch, Chanont Sricharoen, Sarah Lee, Caitlin Reice, Paul Rice, Sophie Kronick, Kim A. Medley, and Kasey D. Fowler-Finn. 2021. “Sex-Specific Ornament Evolution Is a Consistent Feature of Climatic Adaptation across Space and Time in Dragonflies.” PNAS 118 (28): e2101458118. https://doi.org/10.1073/pnas.210145818.
National Public Radio. 2023. “Forest Lizards Have Genetically Morphed to Survive Life in the City, Researchers Say.” National Public Radio (NPR), January 10. Accessed February 19, 2023. https://www.npr.org/2023/01/10/1148150056/forest-lizards-genetically-morphed-cities.
National Science Foundation. 2023. “Urban Lizards Share Genomic Markers Not Found in Forest-Dwellers.” National Science Foundation, February 7. Accessed May 25, 2023. https://beta.nsf.gov/news/urban-lizards-share-genomic-markers-not-found.
Novembre, John, Alison P. Galvani, and Montgomery Slatkin. 2005. “The Geographic Spread of the CCR5 Δ32 HIV-Resistance Allele.” PLoS Biology 3 (11): e339.
Pamuk, Şevket. 2007. “The Black Death and the Origins of the ‘Great Divergence’ across Europe, 1300–1600.” European Review of Economic History 11 (3): 289–317.
Paterlini, Marta. 2007. “There Shall Be Order: The Legacy of Linnaeus in the Age of Molecular Biology.” EMBO Reports 8 (9): 814–816. https://doi.org/10.1038/sj.embor.7401061.
Pew Research Center. 2019. “Darwin in America: The Evolution Debate in the United States.” Pew Research Center, February 6, 2019. Accessed June 13, 2023. https://www.pewresearch.org/religion/2019/02/06/darwin-in-america-2/.
Pew Research Center. 2014. “Fighting Over Darwin, State by State.” Pew Research Center, February 4, 2009; updated February 3, 2014. Accessed May 25, 2022. https://www.pewresearch.org/religion/2009/02/04/fighting-over-darwin-state-by-state/.
Plutynski, Anya. 2009. “The Modern Synthesis.” Routledge Encyclopedia of Philosophy, November 15 (updated December 14, 2009). Accessed November 27, 2022. https://philsci-archive.pitt.edu/15335/.
Plutzer, Eric, Glenn Branch, and Ann Reid. 2020. “Teaching Evolution in the U.S. Public Schools: A Continuing Challenge.” Evolution: Education and Outreach13: Article 14. https://evolution-outreach.biomedcentral.com/articles/10.1186/s12052-020-00126-8.
Proceedings of the National Academy of Sciences (PNAS). 2023. “How Lizards Adapt to Urban Living.” Science Sessions Podcast, February 13, . Accessed February 19, 2023. https://www.pnas.org/post/podcast/lizards-adapt-urban-living.
Public Broadcasting System. 2001. “Georges Cuvier.” WGBH Evolution Library. Accessed May 26, 2022. https://www.pbs.org/wgbh/evolution/library/02/1/l_021_01.html# :~:text=With%20elegant%20studies%20of%20the,as%20incontrovertible%20proof%20of %20extinctions.
Ray, John. 1686-1704. Historia plantarum. London: Clark. Volume 1 (1686), Volume II (1688), Volume III (1704).
Richards, Richard A. 1998. “Darwin, Domestic Breeding, and Artificial Selection.” Endeavour 22 (3): 106–109.
Solloch, Ute V., Kathrin Lang, Vinzenz Lange, and Irena Böhme. 2017. “Frequencies of Gene Variant CCR5-Δ32 in 87 Countries Based on Next-Generation Sequencing of 1.3 Million Individuals Sampled from 3 National DKMS Donor Centers.” Human Immunology 78 (11–12): 701-717.
Stewart, Thomas A., Justin B. Lemberg, Ailis Daly, Edward B. Daeschler, and Neil H. Shubin. 2022. “A New Epistostegalian from the Late Devonian of the Canadian Arctic.” Nature 608 (7923): 563–568.
Tasci, Ufuk Necat. 2020. “How a 10-Century Muslim Physicist Discovered How Humans See.” TRT World, May 25. Accessed May 18, 2022. https://www.trtworld.com/magazine/how-a-10th-century-muslim-physicist-discovered-how-humans-see-36620.
Tbakhi, Abdelghani, and Samir S. Amr. 2007. “Ibn Al-Haytham: Father of Modern Optics.” Annals of Saudi Medicine 27 (6): 464–467. UNAIDS. 2021. “Global HIV & AIDS Statistics—Fact Sheet.” https://www.unaids.org/en/resources/fact-sheet.
UNESCO.org. 2015. “International Year of Light: Ibn al Haytham, Pioneer of Modern Optics celebrated at UNESCO.” UNESCO, September 8. Accessed May 18, 2022. https://www.unesco.org/en/articles/international-year-light-ibn-al-haytham-pioneer-modern-optics-celebrated-unesco.
University of California Berkeley Museum of Paleontology. N.d. “The History of Evolutionary Thought—Uniformitarianism: Charles Lyell.” Understanding Evolution website. Accessed February 13, 2023. https://evolution.berkeley.edu/the-history-of-evolutionary-thought/1800s/uniformitarianism-charles-lyell/.
University of Cambridge. N.d. “Darwin and Dogs.” The Darwin Correspondence Project website. Accessed February 17, 2023. https://www.darwinproject.ac.uk/commentary/curious/darwin-and-dogs.
Urbach, Peter Michael, Anthony M. Quinton, and Kathleen Marguerite Lea. “Francis Bacon: British Author, Philosopher, and Statesman.” Encyclopaedia Britannica. Last updated May 12, 2023. https://www.britannica.com/biography/Francis-Bacon-Viscount-Saint-Alban.
Ward, Peter. 2018. Lamarck’s Revenge: How Epigenetics Is Revolutionizing Our Understanding of Evolution’s Past and Present. New York: Bloomsbury.
Weismann, August. 1892. Das Keimplasma: Eine Theorie der Vererbung (The Germ Plasm: a Theory of Inheritance). Jena (Germany): Fischer.
Weismann, August. 1889. Translations. Essays upon Heredity. Oxford: Clarendon. Accessed November 27, 2022. E-copy available at https://www.esp.org/books/weismann/essays/facsimile/.
Winchell, Kristen M., Shane C. Campbell-Staton, Jonathan B. Losos, and Anthony Geneva. 2023. “Genome-Wide Parallelism Underlies Contemporary Adaptation In Urban Lizards.” PNAS 120 (3): e2216789120. https://doi.org/10.1073/pnas.2216789120.
Wood, Bernard. 2005. Human Evolution: A Very Short Introduction. Oxford: Oxford University Press.
Zou, Yawen. 2015. “The Germ-Plasm: a Theory of Heredity (1893), by August Weismann.” Embryo Project Encyclopedia, January 26. Accessed February 18, 2023. https://embryo.asu.edu/handle/10776/8284.
Hayley Mann, M.A., Binghamton University
Student contributors for this chapter: Emma Costa, Shima Gahima, Will Lefebvre, Audrey Chékinaël
This chapter is a revision from "Chapter 3: Molecular Biology and Genetics" by Hayley Mann, Xazmin Lowman, and Malaina Gaddis. In Explorations: An Open Invitation to Biological Anthropology, first edition, edited by Beth Shook, Katie Nelson, Kelsie Aguilera, and Lara Braff, which is licensed under CC BY-NC 4.0.
Learning Objectives
- Explain and identify the purpose of both DNA replication and the cell cycle.
- Identify key differences between mitosis and meiosis.
- Outline the process of protein synthesis, including transcription and translation.
- Use principles of Mendelian inheritance to predict genotypes and phenotypes of future generations.
- Explain complexities surrounding patterns of genetic inheritance and polygenic traits.
- Discuss challenges to and bioethical concerns of genetic testing.
I [Hayley Mann] started my Bachelor’s degree in 2003, which was the same year the Human Genome Project released its first draft sequence. I initially declared a genetics major because I thought it sounded cool. However, upon taking an actual class, I discovered that genetics was challenging. In addition to my genetics major, I signed up for biological anthropology classes and soon learned that anthropology could bring all those molecular lessons to life. For instance, we are composed of cells, proteins, nucleic acids, carbohydrates, and lipids. Anthropologists often include these molecules in their studies to identify how humans vary; if there are meaningful differences, they propose theories to explain them. Anthropologists study biomolecules in both living and ancient individuals. Ancient biomolecules can also be found on artifacts such as stone tools and cooking vessels. Over the years, scientific techniques for studying organic molecules have improved, which has unlocked new insights into the deep human past.
Cells and Molecules
Molecules of Life
All organisms are composed of four basic types of molecules that are essential for cell structure and function: proteins, lipids, carbohydrates, and nucleic acids (Figure 3.1). Proteins are crucial for cell shape and nearly all cellular tasks, including receiving signals from outside the cell and mobilizing intra-cellular responses. Lipids are a class of organic compounds that include fats, oils, and hormones. Carbohydrates are sugar molecules and serve as energy to cells in the form of glucose. Lastly, nucleic acids, including deoxyribonucleic acid (DNA), carry genetic information about a living organism.
|
Molecule |
Definition |
Example |
|
Proteins |
Composed of one or more long chains of amino acids (i.e., basic units of protein) Often folded into complex 3D shapes that relate to function Proteins interact with other types of proteins and molecules |
Proteins come in different categories including structural (e.g., collagen, keratin, lactase, hemoglobin, cell membrane proteins), defense proteins (e.g, antibodies), enzymes (e.g., lactase), hormones (e.g., insulin), and motor proteins (e.g., actin) |
|
Lipids |
Insoluble in water due to hydrophilic (water-loving) head and a hydrophobic (water-repelling) tail |
Fats, such as triglycerides, store energy for your body Steroid hormones (e.g., estrogen and testosterone) act as chemical messengers to communicate between cells and tissues, as well as biochemical pathways inside of the cell |
|
Carbohydrates |
Large group of organic molecules that are composed of carbon and hydrogen atoms |
Starches and sugars, including blood glucose, provide cells with energy |
|
Nucleic Acids |
Carries the genetic information of an organism |
DNA RNA |
Cells
In 1665, Robert Hooke observed slices of plant cork using a microscope. Hooke noted that the microscopic plant structures he saw resembled cella, meaning “a small room” in Latin. Approximately two centuries later, biologists recognized the cell as being the most fundamental unit of life and that all life is composed of cells. Cellular organisms can be characterized as two main cell types: prokaryotes and eukaryotes (Figure 3.2).
Prokaryotes include bacteria and archaea, and they are composed of a single cell. Additionally, their DNA and organelles are not surrounded by individual membranes. Thus, no compartments separate their DNA from the rest of the cell (see Figure 3.2). It is well known that some bacteria can cause illness in humans. For instance, Escherichia coli (E. coli) and Salmonella contamination can result in food poisoning symptoms. Pneumonia and strep throat are caused by Streptococcal bacteria. Neisseria gonorrhoeae is a sexually transmitted bacterial disease. Although bacteria are commonly associated with illness, not all bacteria are harmful. For example, researchers are studying the relationship between the microbiome and human health. The bacteria that are part of the healthy human microbiome perform beneficial roles, such as digesting food, boosting the immune system, and even making vitamins (e.g., B12 and K).
Eukaryotes can be single-celled or multi-celled in their body composition. In contrast to prokaryotes, eukaryotes possess membranes that surround their DNA and organelles. An example of a single-celled eukaryote is the microscopic algae found in ponds (phytoplankton), which can produce oxygen from the sun. Yeasts are also single-celled, and fungi can be single- or multicellular. Plants and animals are all multicellular.
Although plant and animal cells have a surprising number of similarities, there are some key differences (Figure 3.3). For example, plant cells possess a thick outer cell membrane made of a fibrous carbohydrate called cellulose. Animal and plant cells also have different tissues. For most plants, the outermost layer of cells forms a waxy cuticle that helps to protect the cells and to prevent water loss. Humans have skin, which is the outermost cell layer that is predominantly composed of a tough protein called keratin. Overall, humans have a diversity of tissue types (e.g., cartilage, brain, and heart).
Animal Cell Organelles
An animal cell is surrounded by a double membrane called the phospholipid bilayer (Figure 3.4). A closer look reveals that this protective barrier is made of lipids and proteins that provide structure and function for cellular activities, such as regulating the passage of molecules and ions (e.g., H2O and sodium) into and out of the cell. Cytoplasm is the jelly-like matrix inside of the cell membrane. Part of the cytoplasm comprises organelles, which perform different specialized tasks for the cell (Figure 3.5). An example of an organelle is the nucleus, where the cell’s DNA is located.
Another organelle is the mitochondrion. Mitochondria are often referred to as “powerhouse centers” because they produce energy for the cell in the form of adenosine triphosphate (ATP). Depending on the species and tissue type, multicellular eukaryotes can have hundreds to thousands of mitochondria in each of their cells. Scientists have determined that mitochondria were once symbiotic prokaryotic organisms (i.e., helpful bacteria) that transformed into cellular organelles over time. This evolutionary explanation helps explain why mitochondria also have their own DNA, called mitochondrial DNA (mtDNA). All organelles have important physiological functions and disease can occur when organelles do not perform their role optimally. Figure 3.6 lists other organelles found in the cell and their specialized cellular roles.
|
Cell structure |
Description |
|
Centrioles |
Assist with the organization of mitotic spindles, which extend and contract for the purpose of cellular movement during mitosis and meiosis. |
|
Cytoplasm |
Gelatinous fluid located inside of cell membrane that contains organelles. |
|
Endoplasmic reticulum (ER) |
Continuous membrane with the nucleus that helps transport, synthesize, modify, and fold proteins. Rough ER has embedded ribosomes, whereas smooth ER lacks ribosomes. |
|
Golgi body |
Layers of flattened sacs that receive and transmit messages from the ER to secrete and transport proteins within the cell. |
|
Lysosome |
Located in the cytoplasm; contains enzymes to degrade cellular components. |
|
Microtubule |
Involved with cellular movement including intracellular transport and cell division. |
|
Mitochondrion |
Responsible for cellular respiration, where energy is produced by converting nutrients into ATP. |
|
Nucleolus |
Resides inside of the nucleus and is the site of ribosomal RNA (rRNA) transcription, processing, and assembly. |
|
Nucleopore |
Pores in the nuclear envelope that are selectively permeable. |
|
Nucleus |
Contains the cell’s DNA and is surrounded by the nuclear envelope. |
|
Ribosome |
Located in the cytoplasm and also the membrane of the rough endoplasmic reticulum. Messenger RNA (mRNA) binds to ribosomes and proteins are synthesized. |
Introduction to Genetics
Genetics is the study of heredity. Biological parents pass down their genetic traits to their offspring. Although children resemble their parents, genetic traits often vary in appearance or molecular function. For example, two parents with normal color vision can sometimes produce a son with red-green colorblindness. Molecular geneticists study the biological mechanisms responsible for creating variation between individuals, such as DNA mutations (see Chapter 4), cell division, and genetic regulation.
Molecular anthropologists use genetic data to test anthropological questions. Some of these anthropologists utilize ancient DNA (aDNA), which is DNA that is extracted from anything once living, including human, animal, and plant remains. Over time, DNA becomes degraded (i.e., less intact), but specialized laboratory techniques can make copies of short degraded aDNA segments, which can then be reassembled to provide more complete DNA information.
DNA Structure
The discovery, in 1953, of the molecular structure of deoxyribonucleic acid (DNA) was one of the greatest scientific achievements of all time. Using X-ray crystallography, Rosalind Franklin (Figure 3.7) provided an image that clearly showed the double helix shape of DNA. Due to controversy, Franklin’s colleagues received more recognition for the DNA discovery. In 1962, Watson, Crick, and Wilkins won the Nobel Prize, while Franklin, who had died in 1958, was not honoured. Today, her vital contributions and scientific skill are widely recognized.
The double helix shape of DNA can be described as a twisted ladder (Figure 3.8). More specifically, DNA is a double-stranded molecule with its two strands oriented in opposite directions (i.e., antiparallel). Each strand is composed of nucleotides with a sugar phosphate backbone. There are four different types of DNA nucleotides: adenine (A), thymine (T), cytosine (C), and guanine (G). The two DNA strands are held together by nucleotide base pairs, which have chemical bonding rules. The complementary base-pairing rules are as follows: A and T bond with each other, while C and G form a bond. The chemical bonds between A-T and C-G are formed by “weak” hydrogen atom interactions, which means the two strands can be easily separated. A DNA sequence is the order of nucleotide bases (A, T, G, C) along only one DNA strand. If one DNA strand has the sequence CATGCT, then the other strand will have a complementary sequence GTACGA. This is an example of a short DNA sequence. In reality, there are approximately three billion DNA base pairs in human cells.
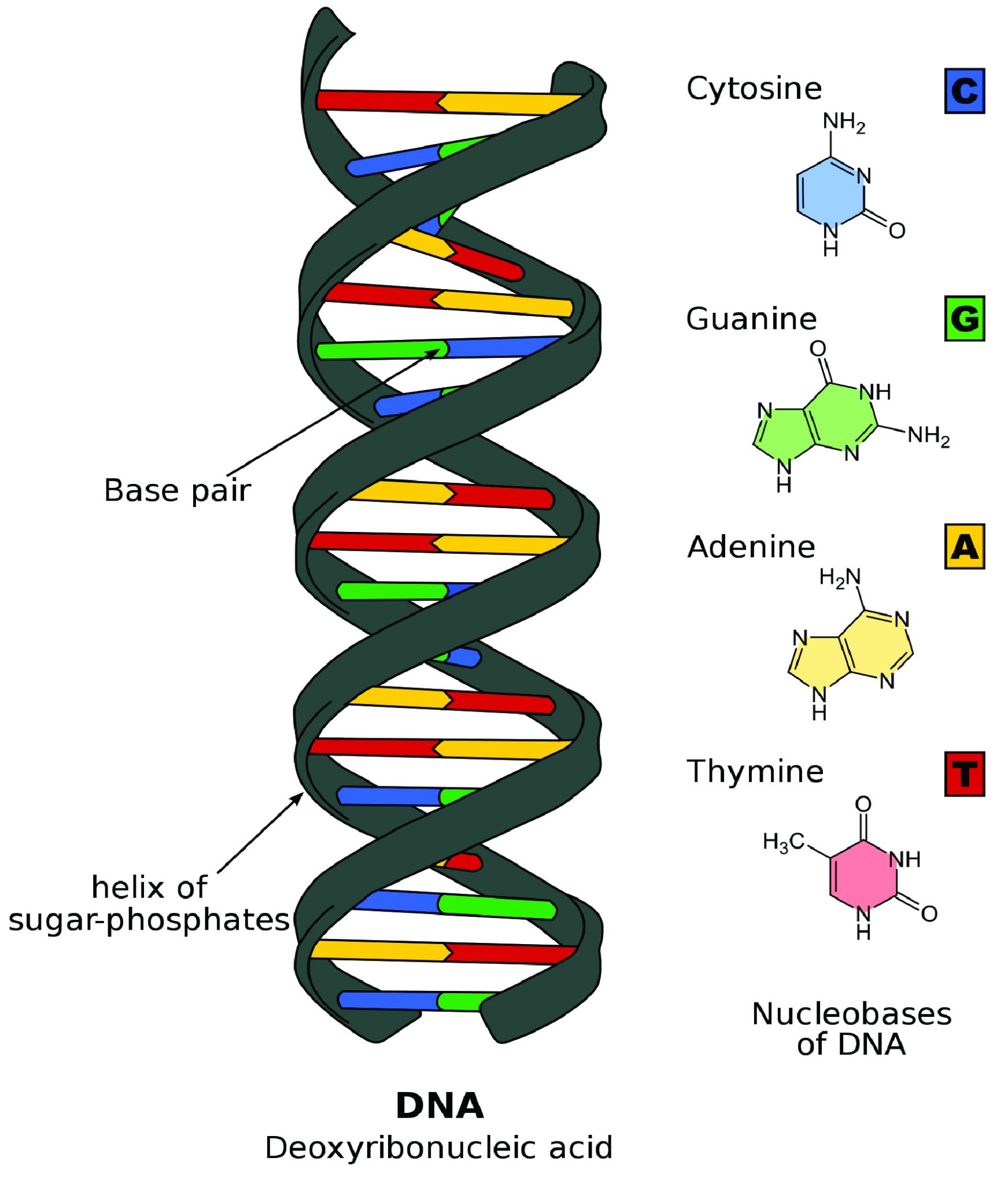
DNA Is Highly Organized within the Nucleus
If you removed the DNA from a single human cell and stretched it out completely, it would measure approximately two meters (about 6.5 feet). Therefore, DNA molecules must be compactly organized in the nucleus. To achieve this, the double helix configuration of DNA undergoes coiling. An analogy would be twisting a string until coils are formed and then continuing to twist so that secondary coils are formed, and so on. To assist with coiling, DNA is first wrapped around proteins called histones. This creates a complex called chromatin, which resembles “beads on a string” (Figure 3.9). Next, chromatin is further coiled into a chromosome. Another important feature of DNA is that chromosomes can be altered from tightly coiled (chromatin) to loosely coiled (euchromatin). Most of the time, chromosomes in the nucleus remain in a euchromatin state so that DNA sequences are accessible for regulatory processes to occur.
Human body cells typically have 23 pairs of chromosomes, for a total of 46 chromosomes in each cell’s nucleus. An interesting fact is that the number of chromosomes an organism possesses varies by species, and this figure is not dependent upon the size or complexity of the organism. For instance, chimpanzees have a total of 48 chromosomes, while hermit crabs have 254. Chromosomes also have a distinct physical structure, including centromeres (the “center”) and telomeres (the ends) (Figure 3.10). Because of the centromeric region, chromosomes are described as having two different “arms,” where one arm is long and the other is shorter. Centromeres play an important role during cell division, which will be discussed in the next section. Telomeres are located at the ends of chromosomes; they help protect the chromosomes from degradation after every round of cell division.
Special Topic: First Nation Immunity and European Diseases—A Study of Ancient DNA

Beginning in the early fifteenth century, First Nations progressively suffered from high mortality rates as the result of colonization from foreign powers. European-borne diseases such as measles, tuberculosis, influenza, and smallpox are largely responsible for the population collapse of Indigenous peoples in the Americas. Many Europeans who immigrated to the Americas had lived in large sedentary populations, which also included coexisting with domestic animals and pests. Although a few prehistoric Indigenous populations can be characterized as large agricultural societies (especially in Mesoamerica), their overall culture, community lifestyle, and subsistence practices were markedly different from that of Europeans. Therefore, because they did not share the same urban living environments as Europeans, it is believed that Indigenous peoples were susceptible to many European diseases.
In 2016, a Nature article published by John Lindo and colleagues was the first to investigate whether pre-contact Indigenous peoples possessed a genetic susceptibility to European diseases. Their study included Tsimshians, a First Nation community from British Columbia (Figure 3.11a-b). DNA from both present-day and ancient individuals (who lived between 500 and 6,000 years ago) was analyzed. The research team discovered that a change occurred in the HLA-DQA1 gene, which is a member of the major histocompatibility complex (MHC) immune system molecules. MHC molecules are responsible for detecting and triggering an immune response against pathogens. Lindo and colleagues (2016) concluded that HLA-DQA1 gene helped Indigenous peoples adapt to their local environmental ecology. However, when European-borne epidemics occurred in the Northwest during the 1800s, a certain HLA-DQA1 DNA sequence variant (allele) associated with ancient Tsimshian immunity was no longer adaptive. As the result of past selective pressures from European diseases, present-day Tsimshians have different HLA-DQA1 allele frequencies. The precise role that HLA-DQA1 plays in immune adaptation requires further investigation. But overall, this study serves as an example of how studying ancient DNA from the remains of deceased individuals can help provide insight into living human populations and historical events.
DNA Replication and Cell Division
For life to continue and flourish, cells must be able to divide. Tissue growth and cellular damage repair are also necessary to maintain an organism throughout its life. All these rely on the dynamic processes of DNA replication and the cell cycle. The mechanisms highlighted in this section are tightly regulated and represent only part of the life cycle of a cell.
DNA Replication
DNA replication is the process by which new DNA is copied from an original DNA template. It is one phase of the highly coordinated cell cycle, and it requires a variety of enzymes with special functions. The creation of a complementary DNA strand from a template strand is described as semi-conservative replication. The result of semi-conservative replication is two separate double-stranded DNA molecules, each of which is composed of an original “parent” template strand and a newly synthesized “daughter” DNA strand.
DNA replication progresses in three steps referred to as initiation, elongation, and termination. During initiation, enzymes are recruited to specific sites along the DNA sequence (Figure 3.12). For example, an initiator enzyme, called helicase, “unwinds” DNA by breaking the hydrogen bonds between the two parent strands. The unraveling of the helix into two separated strands exposes the strands and creates a fork, which is the active site of DNA replication.
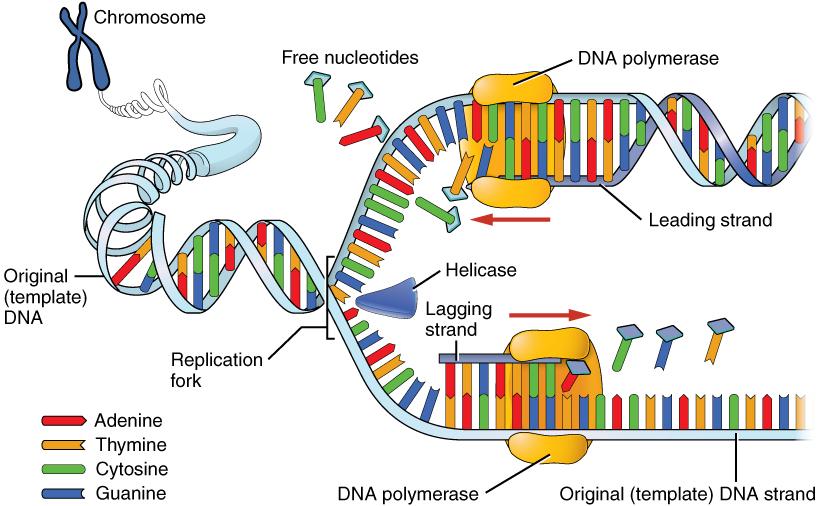
Elongation is the assembly of new DNA daughter strands from the exposed original parent strands. The two parent strands can further be classified as leading strand or lagging strand and are distinguished by the direction of replication. Enzymes called DNA polymerases read parent template strands in a specific direction. Complementary nucleotides are added, and the newly formed daughter strands will grow. On the leading parent strand, a DNA polymerase will create one continuous strand. The lagging parent strand is created in several disconnected sections and other enzymes fill in the missing nucleotide gaps between these sections.
Finally, termination refers to the end of DNA replication activity. It is signaled by a stop sequence in the DNA that is recognized by machinery at the replication fork. The end result of DNA replication is that the number of chromosomes are doubled so that the cell can divide into two.
DNA Mutations
DNA replication should result in the creation of two identical DNA nucleotide sequences. However, although DNA polymerases are quite precise during DNA replication, copying mistakes are estimated to occur every 107 DNA nucleotides. Variation from the original DNA sequence is known as a mutation (Refer to Chapter 4). Briefly, mutations can result in single-nucleotide changes, as well as the insertion or deletion of nucleotides and repeated sequences. Depending on where they occur in the genome, mutations can be deleterious (harmful). For example, mutations may occur in regions that control cell cycle regulation, which can result in cancer (see Special Topic: The Cell Cycle and Immortality of Cancer Cells). Many other types of mutations, however, are not harmful to an organism.
Regardless of their effect, the cell attempts to reduce the frequency of mutations that occur during DNA replication. To accomplish this, there are polymerases with proofreading capacities that can identify and correct mismatched nucleotides. These safeguards reduce the frequency of DNA mutations so that they only occur every 109 nucleotides.
Mitotic Cell Division
There are two types of cells in the body: germ cells (sperm and egg) and somatic cells. The body and its various tissues comprises somatic cells. Organisms that contain two sets of chromosomes in their somatic cells are called diploid organisms. Humans have 46 chromosomes and they are diploid because they inherit one set of chromosomes (n = 23) from each parent. As a result, they have 23 matching pairs of chromosomes, which are known as homologous chromosomes. As seen in Figure 3.13, homologous chromosome pairs vary in size and are generally numbered from largest (chromosome 1) to smallest (chromosome 22) with the exception of the 23rd pair, which is made up of the sex chromosomes (X and Y). Typically, the female sex is XX and the male sex is XY. Individuals inherit an X chromosome from their chromosomal mother and an X or Y from their chromosomal father.
To grow and repair tissues, somatic cells must divide. As discussed previously, for cell division to occur, a cell must first replicate its genetic material. During DNA replication, each chromosome produces double the amount of genetic information. The duplicated arms of chromosomes are known as sister chromatids, and they are attached at the centromeric region. To elaborate, the number of chromosomes stays the same (n = 46); however, the amount of genetic material is doubled in the cell as the result of replication.
Mitosis is the process of somatic cell division that gives rise to two diploid daughter cells (Figure 3.14). Once DNA and other organelles in the cell have finished replication, mitotic spindle fibers physically align each chromosome at the center of the cell. Next, the spindle fibers divide the sister chromatids and move each one to opposite sides of the cell. At this phase, there are 46 chromosomes on each side of a human cell. The cell can now divide into two fully separated daughter cells.
Meiotic Cell Division
Gametogenesis is the production of gametes (sperm and egg cells); it involves two rounds of cell division called meiosis. Similar to mitosis, the parent cell in meiosis is diploid. However, meiosis has a few key differences, including the number of daughter cells produced (four cells, which require two rounds of cell division to produce) and the number of chromosomes each daughter cell has (see Figure 3.14).
During the first round of division (known as meiosis I), each chromosome (n = 46) replicates its DNA so that sister chromatids are formed. Next, with the help of spindle fibers, homologous chromosomes align near the center of the cell and sister chromatids physically swap genetic material. In other words, the sister chromatids of matching chromosomes cross over with each other at matching DNA nucleotide positions. The occurrence of homologous chromosomes crossing over, swapping DNA, and then rejoining segments is called genetic recombination. The “genetic shuffling” that occurs in gametes increases organismal genetic diversity by creating new combinations of genes on chromosomes that are different from the parent cell. Genetic mutations can also arise during recombination. For example, there may be an unequal swapping of genetic material that occurs between the two sister chromatids, which can result in deletions or duplications of DNA nucleotides. Once genetic recombination is complete, homologous chromosomes are separated and two daughter cells are formed.
The daughter cells after the first round of meiosis are haploid, meaning they only have one set of chromosomes (n = 23). During the second round of cell division (known as meiosis II), sister chromatids are separated and two additional haploid daughter cells are formed. Therefore, the four resulting daughter cells have one set of chromosomes (n = 23), and they also have a genetic composition that is not identical to the parent cells nor to each other.
Although both sperm and egg gamete production undergo meiosis, they differ in the final number of viable daughter cells. In the case of spermatogenesis, four mature sperm cells are produced. Although four egg cells are also produced in oogenesis, only one of these egg cells will result in an ovum (mature egg). During fertilization, an egg cell and sperm cell fuse, which creates a diploid cell that develops into an embryo. The ovum also provides the cellular organelles necessary for embryonic cell division. This includes mitochondria, which is why humans, and most other multicellular eukaryotes, have the same mtDNA sequence as their mothers.
Chromosomal Disorders: Aneuploidies
During mitosis or meiosis, entire deletions or duplications of chromosomes can occur due to error. For example, homologous chromosomes may fail to separate properly, so one daughter cell may end up with an extra chromosome while the other daughter cell has one less. Cells with an unexpected (or abnormal) number of chromosomes are known as aneuploid. Adult or embryonic cells can be tested for chromosome number (karyotyping). Aneuploid cells are typically detrimental to a dividing cell or developing embryo, which can lead to a loss of pregnancy. However, the occurrence of individuals being born with three copies of the 21st chromosome is relatively common; this genetic condition is known as Down Syndrome. Moreover, individuals can also be born with aneuploid sex chromosome conditions such as XXY, XXX, and XO (referring to only one X chromosome).
Special Topic: The Cell Cycle and Immortality of Cancer Cells
DNA replication is part of a series of preparatory phases that a cell undergoes prior to cell division, collectively known as interphase (Figure 3.15). During interphase, the cell not only doubles its chromosomes through DNA replication, but it also increases its metabolic capacity to provide energy for growth and division. Transition into each phase of the cell cycle is tightly controlled by proteins that serve as checkpoints. If a cell fails to pass a checkpoint, then DNA replication and/or cell division will not continue. Some of the reasons why a cell may fail at a checkpoint is DNA damage, lack of nutrients to continue the process, or insufficient size. In turn, a cell may undergo apoptosis, which is a mechanism for cell death.
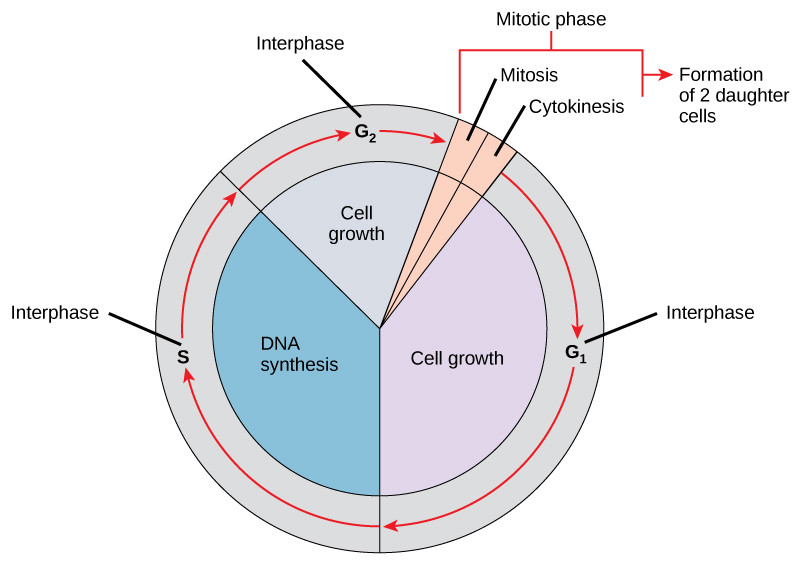
Unchecked cellular growth is a distinguishing hallmark of cancer. In other words, as cancer cells grow and proliferate, they acquire the capacity to avoid death and replicate indefinitely. This uncontrolled and continuous cell division is also known as “immortality.” As previously mentioned, most cells lose the ability to divide due to shortening of telomeres on the ends of chromosomes over time. One way in which cancer cells retain replicative immortality is that the length of their telomeres is continuously protected. Chemotherapy, often used to treat cancer, targets the cell cycle (especially cell division) to halt the propagation of genetically abnormal cells. Another therapeutic approach that continues to be investigated is targeting telomere activity to stop the division of cancer cells.
Researchers have exploited the immortality of cancer cells for molecular research. The oldest immortal cell line is HeLa cells (Figure 3.16), which were harvested from Henrietta Lacks, an African American woman diagnosed with cervical cancer in 1955. At that time, extracted cells frequently died during experiments, but surprisingly HeLa cells continued to replicate. Propagation of Lacks’s cell line has significantly contributed to medical research, including contributing to ongoing cancer research and helping to test the polio vaccine in the 1950s. However, Lacks had not given her consent for her tumor biopsy to be used in cell culture research. Moreover, her family was unaware of the extraction and remarkable application of her cells for two decades. The history of HeLa cell origin was first revealed in 1976. The controversy voiced by the Lacks family was included in an extensive account of HeLa cells published in Rebecca Skloot’s 2010 book, The Immortal Life of Henrietta Lacks. A film based on the book was also released in 2017 (Wolfe 2017).
Protein Synthesis
At the beginning of the chapter, we defined proteins as strings of amino acids that fold into complex 3-D shapes. There are 20 standard amino acids that can be strung together in different combinations in humans, and the result is that proteins can perform an impressive amount of different functions. For instance, muscle fibers are proteins that help facilitate movement. A special class of proteins (immunoglobulins) help protect the organism by detecting disease-causing pathogens in the body. Protein hormones, such as insulin, help regulate physiological activity. Blood hemoglobin is a protein that transports oxygen throughout the body. Enzymes are also proteins, and they are catalysts for biochemical reactions that occur in the cell (e.g., metabolism). Larger-scale protein structures can be visibly seen as physical features of an organism (e.g., hair and nails).
Transcription and Translation
Nucleotides in our DNA provide the coding instructions on how to make proteins. Making proteins, also known as protein synthesis, can be broken down into two main steps referred to as transcription and translation. The purpose of transcription, the first step, is to make an ribonucleic acid (RNA) copy of our genetic code. Although there are many different types of RNA molecules that have a variety of functions within the cell, we will mainly focus on messenger RNA (mRNA). Transcription concludes with the processing (splicing) of the mRNA. The second step, translation, uses mRNA as the instructions for chaining together amino acids into a new protein molecule (Figure 3.17).
Unlike double-stranded DNA, RNA molecules are single-stranded nucleotide sequences (Figure 3.18). Additionally, while DNA contains the nucleotide thymine (T), RNA does not—instead its fourth nucleotide is uracil (U). Uracil is complementary to (or can pair with) adenine (A), while cytosine (C) and guanine (G) continue to be complementary to each other.
For transcription to proceed, a gene must first be turned “on” by the cell. A gene is a segment of DNA that codes for RNA, and genes can vary in length from a few hundred to as many as two million base pairs in length. The double-stranded DNA is then separated, and one side of the DNA is used as a coding template that is read by RNA polymerase. Next, complementary free-floating RNA nucleotides are linked together (Figure 3.19) to form a single-stranded mRNA. For example, if a DNA template is TACGGATGC, then the newly constructed mRNA sequence will be AUGCCUACG.
Genes contain segments called introns and exons. Exons are considered “coding” while introns are considered “noncoding”—meaning the information they contain will not be needed to construct proteins. When a gene is first transcribed into pre-mRNA, introns and exons are both included (Figure 3.20). However, once transcription is finished, introns are removed in a process called splicing. During splicing, a protein/RNA complex attaches itself to the pre-mRNA. Next, introns are removed and the remaining exons are connected, thus creating a shorter mature mRNA that serves as a template for building proteins.
As described above, the result of transcription is a single-stranded mRNA copy of a gene. Translation is the process by which amino acids are chained together to form a new protein. During translation, the mature mRNA is transported outside of the nucleus, where it is bound to a ribosome (Figure 3.21). The nucleotides in the mRNA are read in triplets, which are called codons. Each mRNA codon corresponds to an amino acid, which is carried to the ribosome by a transfer RNA (tRNA). Thus, tRNAs is the link between the mRNA molecule and the growing amino acid chain.
Continuing with our mRNA sequence example from above, the mRNA sequence AUG-CCU-ACG codes for three amino acids. Using a codon table (Figure 3.22), AUG is a codon for methionine (Met), CCU is proline (Pro), and ACG is threonine (Thr). Therefore, the protein sequence is Met-Pro-Thr. Methionine is the most common “start codon” (AUG) for the initiation of protein translation in eukaryotes. As the ribosome moves along the mRNA, the growing amino acid chain exits the ribosome and folds into a protein. When the ribosome reaches a “stop” codon (UAA, UAG, or UGA), the ribosome stops adding any new amino acids, detaches from the mRNA, and the protein is released. Depending upon the amino acid sequence, a linear protein may undergo additional “folding.” The final three-dimensional protein shape is integral to completing a specific structural or functional task.
Dig Deeper: Protein Synthesis
To see protein synthesis in animation, please check out the From DNA to Protein video on YourGenome.org.
Mendelian Genetics

Gregor Johann Mendel (1822–1884) is often described as the “Father of Genetics.” Mendel was a monk who conducted pea plant breeding experiments in a monastery located in the present-day Czech Republic (Figure 3.23). After several years of experiments, Mendel presented his work to a local scientific community in 1865 and published his findings the following year. Although his meticulous effort was notable, the importance of his work was not recognized for another 35 years. One reason for this delay in recognition is that his findings did not agree with the predominant scientific viewpoints on inheritance at the time. For example, it was believed that parental physical traits “blended” together and offspring inherited an intermediate form of that trait. In contrast, Mendel showed that certain pea plant physical traits (e.g., flower color) were passed down separately to the next generation in a statistically predictable manner. Mendel also observed that some parental traits disappeared in offspring but then reappeared in later generations. He explained this occurrence by introducing the concept of “dominant” and “recessive” traits. Mendel established a few fundamental laws of inheritance, and this section reviews some of these concepts. Moreover, the study of traits and diseases that are controlled by a single gene is commonly referred to as Mendelian genetics.
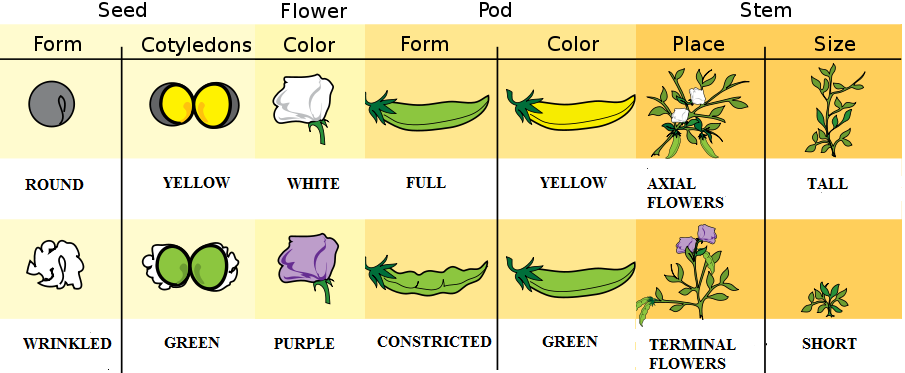
The physical appearance of a trait is called an organism’s phenotype. Figure 3.24 shows pea plant (Pisum sativum) phenotypes that were studied by Mendel, and in each of these cases the physical traits are controlled by a single gene. In the case of Mendelian genetics, a phenotype is determined by an organism’s genotype. A genotype consists of two gene copies, wherein one copy was inherited from each parent. Gene copies are also known as alleles (Figure 3.25), which means they are found in the same gene location on homologous chromosomes. Alleles have a nonidentical DNA sequence, which means their phenotypic effect can be different. In other words, although alleles code for the same trait, different phenotypes can be produced depending on which two alleles (i.e., genotypes) an organism possesses. For example, Mendel’s pea plants all have flowers, but their flower color can be purple or white. Flower color is therefore dependent upon which two color alleles are present in a genotype.
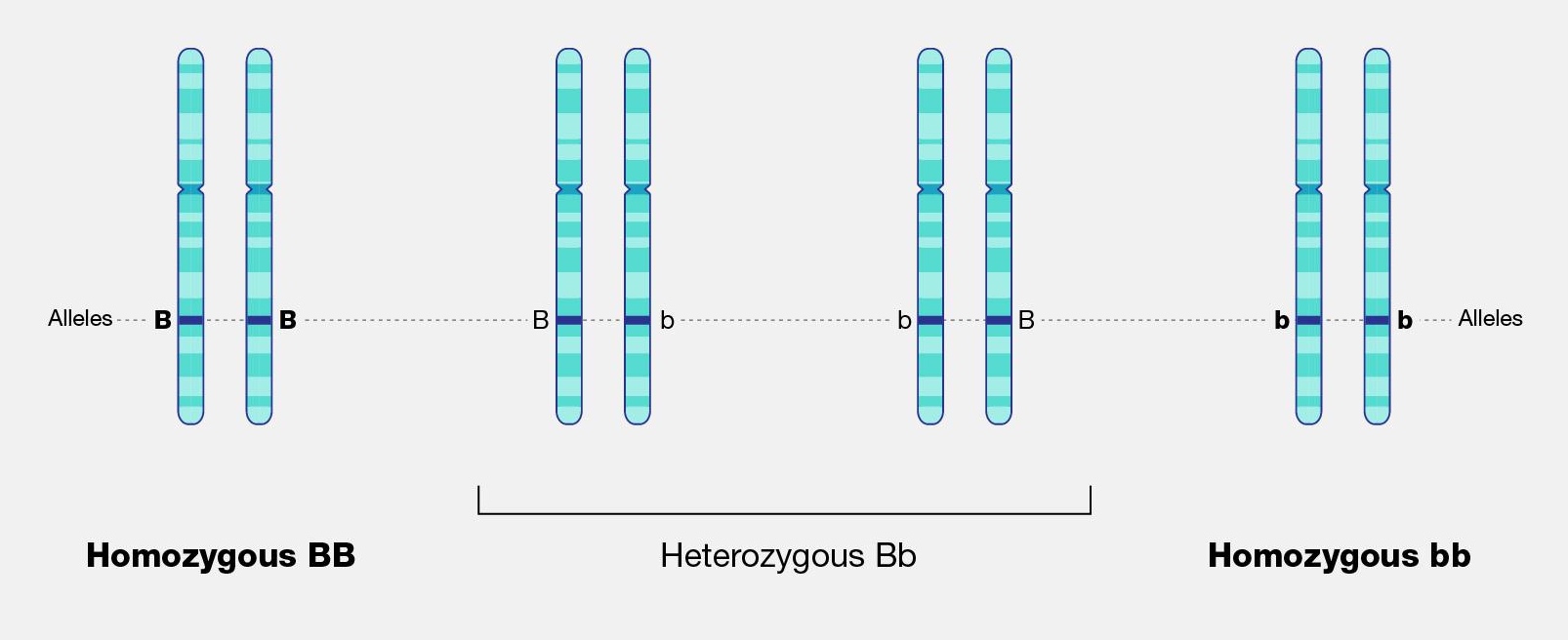
A Punnett square is a diagram that can help visualize Mendelian inheritance patterns. For instance, when parents of known genotypes mate, a Punnett square can help predict the ratio of Mendelian genotypes and phenotypes that their offspring would possess. When discussing genotype, biologists use upper and lower case letters to denote the different allele copies. Figure 3.26 is a Punnett square that includes two heterozygous parents for flower color (Bb). A heterozygous genotype means there are two different alleles for the same gene. Therefore, a pea plant that is heterozygous for flower color has one purple allele and one white allele. When an organism is homozygous for a specific trait, it means their genotype consists of two copies of the same allele. Using the Punnett square example, the two heterozygous pea plant parents can produce offspring with two different homozygous genotypes (BB or bb) or offspring that are heterozygous (Bb).
A pea plant with purple flowers could be heterozygous (Bb) or homozygous (BB). This is because the purple color allele (B) is dominant to the white color allele (b), and therefore it only needs one copy of that allele to phenotypically express purple flowers. Because the white flower allele is recessive, a pea plant must be homozygous for the recessive allele in order to have a white color phenotype (bb). As seen by the Punnett square example (Figure 3.26), three of four offspring will have purple flowers and the other one will have white flowers.
The Law of Segregation was introduced by Mendel to explain why we can predict the ratio of genotypes and phenotypes in offspring. As discussed previously, a parent will have two alleles for a certain gene (with each copy on a different homologous chromosome). The Law of Segregation states that the two copies will be segregated from each other and will each be distributed to their own gamete. We now know that the process where that occurs is meiosis.
Offspring are the products of two gametes combining, which means the offspring inherits one allele from each gamete for most genes. When multiple offspring are produced (like with pea plant breeding), the predicted phenotype ratios are more clearly observed. The pea plants Mendel studied provide a simplistic model to understand single-gene genetics. While many traits anthropologists are interested in have a more complicated inheritance (e.g., are informed by many genes), there are a few known Mendelian traits in humans. Additionally, some human diseases also follow a Mendelian pattern of inheritance (Figure 3.27). Because humans do not have as many offspring as other organisms, we may not recognize Mendelian patterns as easily. However, understanding these principles and being able to calculate the probability that an offspring will have a Mendelian phenotype is still important.
| Mendelian disorder | Gene |
| Alpha Thalassemia | HBA1 |
| Cystic Fibrosis | CFTR |
| Fragile X Syndrome | FMR1 |
| Glucose-6-Phosphate Dehydrogenase Deficiency | G6PD |
| Hemophilia A | F8 |
| Huntington disease | HTT |
| Mitochondrial DNA Depletion Syndrome | TYMP |
| Oculocutaneous Albinism: Type 1 | TYR |
| Polycystic Kidney Disease | PKHD1 |
| Sickle-cell anemia | HBB |
| Spinal Muscular Atrophy: SMN1 Linked | SMN1 |
| Tay-Sachs Disease | HEXA |
| Wilson Disease | ATP7B |
Example of Mendelian Inheritance: The ABO Blood Group System
In 1901, Karl Landsteiner at the University of Vienna published his discovery of ABO blood groups. While conducting blood immunology experiments in which he combined the blood of individuals who possess different blood cell types, he observed an agglutination (clotting) reaction. The presence of agglutination implies there is an incompatible immunological reaction; no agglutination will occur in individuals with the same blood type. This work was clearly important because it resulted in a higher survival rate of patients who received blood transfusions. Blood transfusions from someone with a different type of blood causes agglutinations, and the resulting coagulated blood can not easily pass through blood vessels, resulting in death. Landsteiner received the Nobel Prize (1930) for his discovery and explaination of the ABO blood group system.
Blood cell surface antigens are proteins that coat the surface of red blood cells, and antibodies are specifically “against” or “anti” to the antigens from other blood types. Thus, antibodies are responsible for causing agglutination between incompatible blood types. Understanding the interaction of antigens and antibodies helps to determine ABO compatibility amongst blood donors and recipients. To better comprehend blood phenotypes and ABO compatibility, blood cell antigens and plasma antibodies are presented in Figure 3.28. Individuals that are blood type A have A antigens on the red blood cell surface, and anti-B antibodies, which will bind to B antigens should they come in contact. Alternatively, individuals with blood type B have B antigens and anti-A antibodies. Individuals with blood type AB have both A and B antigens but do not produce antibodies for the ABO system. This does not mean type AB does not have any antibodies present, just that specifically anti-A and anti-B antibodies are not produced. Individuals who are blood type O have nonspecific antigens and produce both anti-A and anti-B antibodies.
Figure 3.29 shows a table of the ABO allele system, which has a Mendelian pattern of inheritance. Both the A and B alleles function as dominant alleles, so the A allele always codes for the A antigen, and the B allele codes for the B antigen. The O allele differs from A and B, because it codes for a nonfunctional antigen protein, which means there is no antigen present on the cell surface of O blood cells. To have blood type O, two copies of the O allele must be inherited, one from each parent, thus the O allele is considered recessive. Therefore, someone who is a heterozygous AO genotype is phenotypically blood type A, and a genotype of BO is blood type B. The ABO blood system also provides an example of codominance, which is when both alleles are observed in the phenotype. This is true for blood type AB: when an individual inherits both the A and B alleles, then both A and B antigens will be present on the cell surface.
Also found on the surface of red blood cells is the rhesus group antigen, known as “Rh factor.” In reality, there are several antigens on red blood cells independent from the ABO blood system, however, the Rh factor is the second most important antigen to consider when determining blood donor and recipient compatibility. Rh antigens must also be considered when a pregnant mother and her baby have incompatible Rh factors. In such cases, a doctor can administer necessary treatment steps to prevent pregnancy complications and hemolytic disease, which is when the mother’s antibodies break down the newborn’s red blood cells.
An individual can possess the Rh antigen (be Rh positive) or lack the Rh antigen (be Rh negative). The Rh factor is controlled by a single gene and is inherited independently of the ABO alleles. Therefore, all blood types can either be positive (O+, A+, B+, AB+) or negative (O-, A-, B-, AB-).
Individuals with O+ red blood cells can donate blood to A+, B+, AB+, and O+ blood type recipients. Because O- individuals do not have AB or Rh antigens, they are compatible with all blood cell types and are referred to as “universal donors.” Individuals that are AB+ are considered to be “universal recipients” because they do not possess antibodies against other blood types.
Mendelian Patterns of Inheritance and Pedigrees
A pedigree can be used to investigate a family’s medical history by determining if a health issue is inheritable and will possibly require medical intervention. A pedigree can also help determine if it is a Mendelian recessive or dominant genetic condition. Figure 3.30 is a pedigree example of a family with Huntington’s disease, which has a Mendelian dominant pattern of inheritance. In a standard pedigree, males are represented by a square and females are represented by a circle. Biological family members are connected to a horizontal line, with biological parents above and offspring below. When an individual is affected with a certain condition, the square or circle is filled in as a solid color. With a dominant condition, at least one of the parents will have the disease and an offspring will have a 50% chance of inheriting the affected chromosome. Therefore, dominant genetic conditions tend to be present in every generation. In the case of Huntington’s, some individuals may not be diagnosed until later in adulthood, so parents may unknowingly pass this dominantly inherited disease to their children.
Because the probability of inheriting a disease-causing recessive allele is more rare, recessive medical conditions can skip generations. Figure 3.31 is an example of a family that carries a recessive cystic fibrosis mutation. A parent that is heterozygous for the cystic fibrosis allele has a 50% chance of passing down their affected chromosome to the next generation. If a child has a recessive disease, then it means both of their parents are carriers (heterozygous) for that condition. In most cases, carriers for recessive conditions show no serious medical symptoms. Individuals whose family have a known medical history for certain conditions sometimes seek family planning services (see the Genetic Testing section).
Pedigrees can also help distinguish if a health issue has either an autosomal or X-linked pattern of inheritance. As previously discussed, there are 23 pairs of chromosomes and 22 of these pairs are known as autosomes. The provided pedigree examples (Figure 3.30–31) are autosomally linked genetic diseases. This means the genes that cause the disease are on one of the chromosomes numbered 1 to 22. The conditions caused by genes located on the X chromosome are referred to as X-linked diseases.
Figure 3.32 depicts a family in which the mother is a carrier for the X-linked recessive disease Duchenne Muscular Dystrophy (DMD). The mother is a carrier for DMD, so daughters and sons will have a 50% chance of inheriting the pathogenic DMD allele. Because females have two X chromosomes, females who inherit only one copy will not have the disease (although in rare cases, female carriers may show some symptoms of the disease). On the other hand, males who inherit a copy of an X-linked pathogenic DMD allele will typically be affected with the condition. Thus, males are more susceptible to X-linked conditions because they only have one X chromosome. Therefore, when evaluating a pedigree, if a higher proportion of males are affected with the disease, this could suggest the disease is X-linked recessive.
Compared to the X chromosome, the Y chromosome is smaller with only a few genes. Y-linked traits are therefore rare and can only be passed from a chromosomal father to a biological XY child.
Other Patterns of Inheritance
Complexity Surrounding Mendelian Inheritance
Pea plant trait genetics are relatively simple compared to what we know about genetic inheritance today. The vast majority of genetically controlled traits are not strictly dominant or recessive, so the relationship among alleles and predicting phenotype is often more complicated. For example, traits that exhibit incomplete dominance occur when a heterozygote exhibits a phenotype that is an intermediate phenotype of both alleles. In snapdragon flowers, the red flower color (R) is dominant and white is recessive (r). Therefore, the homozygous dominant RR is red and homozygous recessive rr is white. However, because the R allele is not completely dominant, the heterozygote Rr is a blend of red and white, which results in a pink flower (Figure 3.33).

An example of incomplete dominance in humans is the enzyme β-hexosaminidase A (Hex A), which is encoded by the gene HEXA. Patients with two dysfunctional HEXA alleles are unable to metabolize a specific lipid-sugar molecule (GM2 ganglioside); because of this, the molecule builds up and causes damage to nerve cells in the brain and spinal cord. This condition is known as Tay-Sachs disease, and it usually appears in infants who are three to six months old. Most children with Tay-Sachs do not live past early childhood. Individuals who are heterozygous for the functional type HEXA allele and one dysfunctional allele have reduced Hex A activity. However, the amount of enzyme activity is still sufficient, so carriers do not exhibit any neurological phenotypes and appear healthy.
Some genes and alleles can also have higher penetrance than others. Penetrance can be defined as the proportion of individuals who have a certain allele and also express an expected phenotype. If a genotype always produces an expected phenotype, then those alleles are said to be fully penetrant. However, in the case of incomplete (or reduced) penetrance, an expected phenotype may not occur even if an individual possesses the alleles that are known to control a trait or cause a disease.
A well-studied example of genetic penetrance is the cancer-related genes BRCA1 and BRCA2. Mutations in these genes can affect crucial processes such as DNA repair, which can lead to breast and ovarian cancers. Although BRCA1 and BRCA2 mutations have an autosomal dominant pattern of inheritance, it does not mean an individual will develop cancer if they inherit a pathogenic allele. Several lifestyle and environmental factors can also influence the risk for developing cancer. Regardless, if a family has a history of certain types of cancers, then it is often recommended that genetic testing be performed for individuals who are at risk. Moreover, publically available genetic testing companies are now offering health reports that include BRCA1 and BRCA2 allele testing (see the Genetic Testing section).
Polygenic Traits
While Mendelian traits tend to be influenced by a single gene, the vast majority of human phenotypes are polygenic traits. The term polygenic means “many genes.” Therefore, a polygenic trait is influenced by many genes that work together to produce the phenotype. Human phenotypes such as hair color, eye color, height, and weight are examples of polygenic traits. Hair color, for example, is largely determined by the type and quantity of a pigment called melanin, which is produced by a specialized cell type within the skin called melanocytes. The quantity and ratio of melanin pigments determine black, brown, blond, and red hair colors. MC1R is a well-studied gene that encodes a protein expressed on the surface of melanocytes that is involved in the production of eumelanin pigment. Typically, people with two functional copies of MC1R have brown hair. People with reduced functioning MC1R allele copies tend to produce pheomelanin, which results in blond or red hair. However, MC1R alleles have variable penetrance, and studies are continually identifying new genes (e.g., TYR, TYRP1, SLC24A5, and KITLG) that also influence hair color. Individuals with two nonfunctioning copies of the gene TYR have a condition called oculocutaneous albinism—their melanocytes are unable to produce melanin so these individuals have white hair, light eyes, and pale skin.
In comparison to Mendelian diseases, complex diseases (e.g., Type II diabetes, coronary heart disease, Alzheimer's, and schizophrenia) are more prevalent in humans. Complex diseases are polygenic, but their development is also influenced by physical, environmental, sociocultural, and individual lifestyle factors. Families can be more predisposed to certain diseases; however, complex diseases often do not have a clear pattern of inheritance.
Although research of complex traits and diseases continue, geneticists may not know all of the genes involved with a given complex disease. Additionally, how much genetic versus nongenetic determinants contribute to a disease phenotype can be difficult to decipher. Therefore, predicting individual medical risk and risk across different human populations is often a significant challenge. For instance, cardiovascular diseases (CVDs) continue to be one of the leading causes of death around the world. Development of CVDs has been linked to nutrient exposure during fetal development, high fat and sedentary lifestyles, drug usage, adverse socioeconomic conditions, and various genes. Human environments are diverse, and public health research including the field of Human Biology can help identify risk factors and behaviors associated with chronic diseases. Large-scale clinical genetic studies with powerful bioinformatic approaches can also help elucidate some of these complex relationships.
Genomics and Epigenetics
A genome is all of the genetic material of an organism. In the case of humans, this includes 46 chromosomes and mtDNA. The human genome contains approximately three billion base pairs of DNA and has regions that are both noncoding and coding. Scientists now estimate that the human genome contains 20,000–25,000 protein-coding genes, with each chromosome containing a few hundred to a few thousand genes. As our knowledge of heredity increases, researchers have begun to realize the importance of epigenetics, or changes in gene expression that do not result in a change of the underlying DNA sequence. Epigenetics research is also crucial for unraveling gene regulation, which involves complex interactions between DNA, RNA, proteins, and the environment.
Genomics
The vast majority of the human genome is noncoding, meaning there are no instructions to make a protein or RNA product in these regions. Historically, noncoding DNA was referred to as “junk DNA” because these vast segments of the genome were thought to be irrelevant and nonfunctional. However, continual improvement of DNA sequencing technology along with worldwide scientific collaborations and consortia have contributed to our increased understanding of how the genome functions. Through these technological advances and collaborations, we have since discovered that many of these noncoding DNA regions are involved in dynamic genetic regulatory processes.
Genomics is a diverse field of molecular biology that focuses on genomic evolution, structure, and function; gene mapping; and genotyping (determining the alleles present). Evolutionary genomics determined that humans share about 98.8% percent of their DNA with chimpanzees. Given the phenotypic differences between humans and chimpanzees, having a DNA sequence difference of 1.2% seems surprising. However, a lot of genomics research is also focused on understanding how noncoding genomic regions influence how individual genes are turned “on” and “off” (i.e., regulated). Therefore, although DNA sequences are identical, regulatory differences in noncoding genetic regions (e.g., promoters) are believed to be largely responsible for the physical differences between humans and chimpanzees.
Further understanding of genomic regulatory elements can lead to new therapies and personalized treatments for a broad range of diseases. For example, targeting the regulatory region of a pathogenic gene to “turn off” its expression can prevent its otherwise harmful effects. Such molecular targeting approaches can be personalized based on an individual’s genetic makeup. Genome-wide association studies (GWAS), which seek to determine genes that are linked to complex traits and diseases, typically require significant computational efforts. This is because millions of DNA sequences must be analyzed and GWAS sometimes include thousands of participants. During the beginning of the genomics field, most of the large-scale genomics studies only included North American, European, and East Asian participants and patients. Researchers are now focusing on increasing ethnic diversity in genomic studies and databases. In turn, accuracy of individual disease risk across all human populations will be improved and more rare disease–causing alleles will be identified.
Epigenetics
All cells within your body have the same copy of DNA. For example, a brain neuron has the same DNA blueprint as does a skin cell on your arm. Although these cells have the same genetic information, they are considered specialized. The reason all cells within the body have the same DNA but different morphologies and functions is that different subsets of genes are turned “on” and “off” within the different cell types. A more precise explanation is that there is differential expression of genes among different cell types. In the case of neuronal cells, a unique subset of genes are active that allow them to grow axons to send and receive messages. This subset of genes will be inactive in non-neuronal cell types such as skin cells. Epigenetics is a branch of genetics that studies how these genes are regulated through mechanisms that do not change the underlying DNA sequence.
The prefix epi- means “on, above, or near,” and epigenetic mechanisms such as DNA methylation and histone modifications occur on, above, or near DNA. The addition of a methyl group (— CH₃) to DNA is known as DNA methylation (Figure 3.34). DNA methylation and other modifications made to the histones around which DNA are wrapped are thought to make chromatin more compact. This DNA is inaccessible to transcription factors and RNA polymerases, thus preventing genes from being turned on (i.e., transcribed). Other histone modifications have the opposite effect by loosening chromatin, which makes genes accessible to transcription factors.
It is important to note that environmental factors can alter DNA methylation and histone modifications and also that these changes can be passed from generation to generation. For example, someone’s epigenetic profile can be altered during a stressful time (e.g., natural disasters, famine, etc.), and those regulatory changes can be inherited by the next generation. Moreover, our epigenetic expression profile changes as we age. For example, certain places in our genome become “hyper” or “hypo” methylated over time. Identical twins also have epigenetic profiles that become more different as they age. Researchers are only beginning to understand the significance of these genome-wide epigenetic changes. Scientists have also discovered that changes in epigenetic modifications can alter gene expression in ways that contribute to diseases. It is also important to note that, unlike DNA mutations (which permanently change the nucleotide sequence), epigenetic changes can be easily reversed. A lot of research now focuses on how drugs can alter or modulate changes in DNA methylation and histone modifications to treat diseases such as cancer.
Environmental Disruptors and Their Impact on Human Reproductive Systems
The National Institute of Environmental Health Sciences (NIEHS) defines endocrine-disrupting chemicals (EDCs) as synthetic or natural compounds that interfere with the body’s hormonal systems. Found in pesticides, plastics, industrial chemicals, and pollutants, EDCs can mimic, block, or alter the natural action of hormones (NIEHS, 2024). Their effects on reproductive health are profound, particularly during critical developmental windows while also affecting subsequent generations through epigenetic changes.
NIEHS declared EDC’s:
| Atrazine | one of the most commonly applied herbicides in the world, often used to control weeds in corn, sorghum, and sugarcane crops. |
| Bisphenol A (BPA) | used to make polycarbonate plastics and epoxy resins. It is used in manufacturing, food packaging, toys, and other applications. BPA resins may be found in the lining of some canned foods and beverages. |
| Dioxins | a byproduct of certain manufacturing processes, such as herbicide production and paper bleaching. They can be released into the air from waste burning and wildfires. |
| Perchlorate | a colorless salt manufactured and used as an industrial chemical to make rockets, explosives, and fireworks, which can be found in some groundwater. |
| Polyfluoroalkyl Substances (PFAS) | a large group of chemicals used widely in industrial applications, such as firefighting foam, nonstick pans, paper, and textile coatings. |
| Phthalates | a large group of compounds used as liquid plasticizers. They are found in hundreds of products including some food packaging, cosmetics, fragrances, children’s toys, and medical device tubing. Cosmetics that may contain phthalates include nail polish, hair spray, aftershave lotion, cleanser, and shampoo. |
| Phytoestorgens | naturally occurring substances with hormone-like activity found in some plants; they may have a similar effect to estrogen produced by the body. Soy foods, for example, contain phytoestrogens. |
| Polybrominated diphenyl ethers (PBDE) | used to make flame retardants for products such as furniture foam and carpet. |
| Polychlorinated biphenyls (PCBs) | used to make electrical equipment, such as transformers, and are in hydraulic fluids, heat transfer fluids, lubricants, and plasticizers. PCBs were mass-produced globally until they were banned in 1979. |
| Triclosan | an ingredient that was previously added to some antimicrobial and personal care products, like liquid body wash and soaps. |
The Male Reproductive System: Vulnerabilities, Epigenetics, and Disruptions
The male reproductive system is highly sensitive to hormonal interference, especially during prenatal and early postnatal development. Over the past 50 years, epidemiological data gathered by the NIEHS has revealed alarming changes: increased cases of prostate and testicular cancers, male-descended testes, and anatomical malformations of male genitalia (Sweeney et al., 2015). These changes are accompanied by a global decline in sperm quality, underscoring the widespread vulnerability of male reproductive health to environmental factors. The testes, as the site of sperm production and testosterone synthesis, are particularly susceptible to EDC interference. Proper testicular development depends on tightly regulated hormonal signalling, which EDCs can disrupt by mimicking or blocking hormones like testosterone and estrogen, leading to improper testicular formation and increased risk of testicular cancer. Prostate development is also a target for EDC interference. African American men, for example, exhibit twice the risk of developing prostate cancer than Caucasian men. This disparity has been attributed to hereditary, lifestyle, and environmental factors, often causing elevated maternal estrogen levels during gestation. This prenatal exposure to EDCs can mimic estrogen and predispose developing prostate tissues to cancerous changes in adulthood (2015).
The Female Reproductive System: Epigenetics and Fertility Challenges
Female fertility relies on a delicate hormonal balance to regulate processes such as ovulation, implantation, and pregnancy. EDCs can disrupt this balance by mimicking, antagonizing, or altering the action of hormones. Their interference contributes to a wide range of reproductive disorders, including early puberty, premature ovarian failure, anovulation, and infertility. Epigenetics plays a central role in female reproductive health. DNA methylation, histone modifications, and ncRNA generation are crucial for regulating ovarian and uterine function; However, EDCs can affect these regulatory mechanisms. An example of this is primordial germ cells (PGCs) in female embryos, which need to undergo extensive epigenetic reprogramming during development (Biswas et al., 2021). This process erases genomic imprinting and reactivates the inactive X chromosome, creating a "blank slate" for the next generation; however, EDCs can disrupt this critical period of epigenetic resetting, leading to long-term consequences for reproductive health.
The ovarian follicle–the functional unit of female reproduction–is particularly vulnerable to these chemicals. Being exposed to EDCs can deplete the pool of these follicles, leading to temporary or permanent infertility (2021). Additionally, EDCs interfere with estrogen receptor function, a crucial regulator of female reproductive processes. These chemicals bind to these receptors, altering the recruitment of enzymes involved in histone modification and chromatin remodelling; this disrupts gene expression patterns critical for ovarian and uterine health. One striking example is diethylstilbestrol (DES), a synthetic estrogen once prescribed to pregnant women (2021). DES exposure has been linked to ovarian cancer in subsequent generations, highlighting the transgenerational effects of EDCs on the female reproductive system. In severe cases, EDCs induce multigenerational reproductive disorders, as observed in studies linking DES to ovarian cancer in the grandchildren of exposed individuals.
Epigenetic Therapy
Heritable Changes and Some Related Drugs
As has been said, epigenetics involves heritable changes in gene expression, without involving DNA alteration. These changes, being heritable and often involving abnormal DNA methylation patterns within the four DNA methyltransferases (DNMTs) or histone modifications in chromatin, can lead to disease development. DNMTs (DNMT1, DNMT2, DNMT3A, and DNMT3B) have functions specific to themselves and are at the core of the DNA methylation process. Regarding the histone modifications mentioned, histones have been recognized to mutate under various mechanisms, such as acetylation, methylation and phosphorylation. The acetylation of histones involves histone acetyltransferases (HATs), which are associated with the activation of gene transcription. This process is reversed by the deacetylation of histones, which is associated with the silencing of gene transcription under histone deacetylases (HDACs). (Peedicayil, 2006)
Epigenetic therapy, with the use of specialized drug developments, aims to correct epigenetic defects, which are reversible under pharmacological intervention, by targeting enzymes such as HATs, HDACs and DNMTs, as well as histone methyltransferases. For instance, certain drugs are being developed as DNMT inhibitors, stopping the methylation of DNA associated with inappropriate transcriptional silencing of genes, and potentially increasing haemoglobin F to help patients affected by sickle cell anemia. These DNMT inhibitor drugs have been classified under three categories based on their structures: nucleoside analogue DNMT inhibitors, non-nucleoside analogue DNMT inhibitors, and antisense oligonucleotides (2006). Nucleoside analogue DNMT inhibitors are analogues of cytosine, the nucleotide affected by methylation from DNMTs, and are incorporated into replicating DNA, replacing cytosine, thus being S-phase-specific drugs. Non-nucleoside analogue DNMT inhibitors are researched to reduce the myelotoxic effects of drugs directly incorporated into the DNA, and are brought into the patient differently. Antisense oligonucleotides are drugs made up of sequences of nucleotides complementary to mRNAs, made to block translation, by acting on the DNMT1 for instance. Additionally, drugs such as HDAC inhibitors help maintain the acetylation of histones, leading to apoptosis, growth arrest or differentiation of tumour cells, giving this drug an anticancer effect, suppressing tumour growth. (2006)
Implications with Cancers
Research published in The Indian Journal for Medical Research has shown that these drugs show promising results in cancer treatment trials involving solid tumours and hematological malignancies. However, they have limitations, for instance, the fact that DNMT and HDAC inhibitors could activate oncogenes due to limited specificity, leading to further tumor progression; or their high myelotoxicity levels, a side effect thought to be due to their incorporation into DNA, and nucleotide analogue inhibitors (2006). Though that is the case, it is important to know that epigenetic drugs alone or in combination with conventional anticancer drugs, may prove to be a significant advance over the use of conventional anticancer drugs, and may also be a way to prevent diseases. Additionally, combination therapy strategies targeting various epigenetic markers, such as DNMTs for cancer-related genes and non-selective HDAC inhibitors, have been shown to yield promising results, simultaneously inducing the expression of tumor suppressor genes and inhibiting the expression of key oncogenes. As recently explored by researchers in Cell Death Discovery, this specific case of combination therapy would synergistically induce gene expression while maintaining the selectivity required to increase targeting of particular tumor types based on gene expression profiles. (Yu et al., 2024)
To date, the majority of cases in which epigenetic defects have led to disease pathogenesis are cancers (Peedicayil, 2006), cancer cells often developing due to uncontrolled cell growth and resistance to cell death mechanisms, made possible with abnormal DNA methylation patterns as well as histone modifications (Yu et al., 2024). Epigenetic alterations have therefore been identified within the core of tumor progression mechanisms in cancer cells, including tumorigenesis, promotion, progression, and recurrence, suggesting epigenetic heterogeneity at the cellular level (2024). Certain drugs have been developed, showing specifically good results for cancer treatments, by inhibiting enzymes such as KMTs and KDMs. These can be added to the growing list of drugs fitting into epigenetic therapy, including DNMT and HDAC inhibitors, as well as combination therapy treatments, for cancer and other diseases.
Purpose of Study and Future Developments
Studying the link between epigenetics and diseases is crucial for multiple reasons, one of which is enabling scientists and researchers to better understand disease mechanisms, detect abnormal epigenetic changes, and, in turn, develop more effective treatments or possibly even prevent diseases from developing in the first place. As previously mentioned, epigenetic therapy has been shown to bring promising results in drug trials surrounding cancer treatments. Still, the range of diseases to be treated with this new pharmacology approach is vast, molecules other than DNMTs and HDACs being related to epigenetic mechanisms within gene expression, such as BET proteins and KDMs, potentially being a source of new medications or treatments (Yu et al., 2024; Peedicayil, 2006). Additionally, by understanding someone's epigenetic profile, a form of personalized “precision medicine” (Yu et al., 2024, p. 8) is developed, offering less toxic and more effective treatments with fewer undesired side effects. Researchers expanding this field of knowledge would be able to understand, in more concrete terms, how external factors are linked to epigenetic changes and, consequently, disease risk, potentially halting disease progression and developing new prevention mechanisms. Personalized medicine combines both genetic and epigenetic data, including gene expression profiles, DNA methylation patterns, histone modification profiles, and identified biomarkers, to create precise disease management and prediction.
It is crucial to keep in mind that diseases like cancer are linked to major causes of morbidity and mortality worldwide, which could be reduced with therapeutic medicine such as epigenetic therapy, aiming to detect cancer biomarkers to improve risk assessment, diagnosis, and targeted treatment interventions, limiting the burden of chronic and life-threatening diseases. With the advancement of epigenetic therapies, new sequencing techniques, as well as AI (2024), have opened avenues to establish precision diagnostics and therapeutics for patients.
With this said, epigenetics is a relatively new area of scientific research. This field has exploded in the last few decades, especially with the advancement of technologies that allow researchers to examine DNA methylation patterns, histone modifications, and non-coding RNA molecules across the genome. While the potential of epigenetics in explaining complex diseases, including those linked to environmental factors such as endocrine-disrupting chemicals (EDCs), is immense, we’ve identified two key challenges. One major limitation is the complexity and variability of epigenetic marks. These modifications can differ significantly across cell types, tissues, and even individuals, making it difficult to generalize findings.
Additionally, epigenetic changes are dynamic and can fluctuate over time, which complicates the task of linking them to specific environmental exposures or health outcomes. Another challenge lies in the transgenerational aspect of epigenetics. While it's clear that epigenetic changes can be passed from one generation to the next, the mechanisms behind this inheritance are not fully understood. It's also difficult to pinpoint exactly when and how these modifications occur in development, especially since environmental exposures may affect individuals at different stages of their life, with varying effects depending on the timing and dose.
Special Topic: Epigenetics and X Chromosome Inactivation

Mary Lyon was a British geneticist who presented a hypothesis for X chromosome inactivation (called the Lyon hypothesis) based on her work and other studies of the day. Females inherit two X chromosomes, one from each parent. Males have one functional X chromosome; however, this does not mean females have more active genes than males. During the genetic embryonic development of many female mammals, one of the X chromosomes is inactivated at random, so females have one functional X chromosome. The process of X chromosome inactivation in females occurs through epigenetic mechanisms, such as DNA methylation and histone modifications. Recent studies have analyzed the role of a long noncoding RNA called X-inactive specific transcript (XIST), which is largely responsible for the random silencing of one of the X chromosomes. The presence of two X chromosomes is the signal for XIST RNA to be expressed so that one X chromosome can be inactivated. However, some cells may have an active paternal X chromosome while other cells may have an active maternal X chromosome. This phenomenon is easily seen in calico and tortoiseshell cats (Figure 3.35). In cats, the gene that controls coat color is found on the X chromosome. During early embryo development, random inactivation of X chromosomes gives rise to populations of cells that express black or orange, which results in the unique coat patterning. Therefore, calico cats are typically always female.
Genetic Testing
To assist with public health efforts, newborn screening for genetic diseases has been available in the United States for over 50 years. One of the first available genetic tests was to confirm a phenylketonuria (PKU) diagnosis in infants, which is easily treatable with a dietary change. Currently, each state decides what genes are included on newborn screening panels and some states even have programs to help with infant medical follow-ups. There are now hundreds of laboratories that provide testing for a few thousand different genes that can inform medical decisions for infants and adults. Moreover, genetic testing has been made available publicly to anyone without the assistance of medical professionals.
Clinical Testing
Clinical genetics tests assist patients with making medically informed decisions about family planning and health. Applications of this technology include assistance with in vitro fertilization (IVF) procedures, embryo genetic screening, and personalized medicine such as matching patients to cancer therapies. To ensure accuracy of patient genetic screening, it is important that all clinical laboratories are regulated. The Clinical Laboratory Improvement Amendments (CLIA) are United States federal standards that all human laboratory testing clinics must follow. A major benefit provided by some clinical genetic testing companies is access to genetic counselors, who have specialized education and training in medical genetics and counseling. For individuals with a family history of genetic disease, a physician may recommend genetic carrier screening to see if there is a risk for passing on a disease to a child. Genetic counselors provide expertise with interpretation of genetic testing results, as well as help guide and support patients when making impactful medical decisions.
Review Questions
- What is the purpose of DNA replication? Explain in a few sentences what happens during DNA replication. When do DNA mutations happen? And how does this create phenotypic variation (i.e., different phenotypes of the same physical trait)?
- Using your own words, what are homologous chromosomes and sister chromatids? What are the key differences between mitosis and meiosis?
- Determine if the pedigree diagram below (Figure 3.41) represents an autosomal dominant, autosomal recessive, or X-linked recessive pattern of inheritance. You should write the genotype (i.e., AA, Aa, or aa) above each square to help you (note: there may sometimes be two possible answers for a square’s genotype). Please also explain why you concluded a particular pattern of inheritance.
- Use base pairing rules to transcribe the following DNA template sequence into mRNA: GTAAAGGTGCTGGCCATC. Next, use the protein codon table (see Figure 3.21) to translate the sequence. In regard to transcription, explain what the significance is of the first and last codon/protein in the sequence.
- In your opinion, what do you think the benefits are of direct-to-consumer (DTC) genetic testing? What are the drawbacks and/or greater ethical concerns? Do you think benefits outweigh concerns?
- Imagine that you submit your DNA sample to a genetic testing company and among the various diseases for which they test, there is an allele that is associated with late-onset Alzheimer’s disease. You have the option to view your Alzheimer’s result or to not view your result. What do you do and why?
Key Terms
Adenosine triphosphate (ATP): A high-energy compound produced by mitochondria that powers cellular processes.
Allele: A nonidentical DNA sequence found in the same gene location on a homologous chromosome, or gene copy, that codes for the same trait but produces a different phenotype.
Amino acids: Organic molecules that are the building blocks of protein. Each of the 20 different amino acids have their own unique chemical property. Amino acids are chained together to form proteins.
Ancient DNA (aDNA): DNA that is extracted from organic remains and that often dates from hundreds to thousands of years ago. Also, aDNA is typically degraded (i.e., damaged) due to exposure to the elements such as heat, acidity, and humidity.
Aneuploid: A cell with an unexpected amount of chromosomes. The loss or gain of chromosomes can occur during mitotic or meiotic division.
Antibodies: Immune-related proteins that can detect and bind to foreign substances in the blood such as pathogens.
Apoptosis: A series of molecular steps that is activated leading to cell death. Apoptosis can be activated when a cell fails checkpoints during the cell cycle; however, cancer cells have the ability to avoid apoptosis.
Autosomal: Refers to a pattern of inheritance in which an allele is located on an autosome (non sex chromosome).
Base pairs: Chemical bonding between nucleotides. In DNA, adenine (A) pairs with thymine (T) and cytosine (C) pairs with guanine (G); in RNA, adenine (A) always pairs with uracil (U).
Carbohydrate: Molecules composed of carbon and hydrogen atoms that can be broken down to supply energy.
Carrier: An individual who has a heterozygous genotype that is typically associated with a disease.
Cell cycle: A cycle the cell undergoes with checkpoints between phases to ensure that DNA replication and cell division occur properly.
Cell surface antigen: A protein that is found on a red blood cell’s surface.
Centromere: A structural feature that is defined as the “center” of a chromosome and that creates two different arm lengths. This term also refers to the region of attachment for microtubules during mitosis and meiosis.
Chromatin: DNA wrapped around histone complexes. During cell division, chromatin becomes a condensed chromosome.
Chromosome: DNA molecule that is wrapped around protein complexes, including histones.
Codominance: The effects of both alleles in a genotype can be seen in the phenotype.
Codons: A sequence that comprises three DNA nucleotides that together code for a protein.
Complex diseases: A category of diseases that are polygenic and are also influenced by environment and lifestyle factors.
Cytoplasm: The “jelly-like” matrix inside of the cell that contains many organelles and other cellular molecules.
Deleterious: A mutation that increases an organism’s susceptibility to disease.
Deoxyribonucleic acid (DNA): A molecule that carries the hereditary information passed down from parents to offspring. DNA can be described as a “double helix”’ shape. It includes two chains of nucleotides held together by hydrogen bonds with a sugar phosphate backbone.
Diploid: Refers to an organism or cell with two sets of chromosomes.
DNA methylation: Methyl groups bind DNA, which modifies the transcriptional activity of a gene by turning it “on” or “off.”
DNA polymerase: Enzyme that adds nucleotides to existing nucleic acid strands during DNA replication. These enzymes can be distinguished by their processivity (e.g., DNA replication).
DNA replication: Cellular process in which DNA is copied and doubled.
DNA sequence: The order of nucleotide bases. A DNA sequence can be short, long, or representative of entire chromosomes or organismal genomes.
Dominant: Refers to an allele for which one copy is sufficient to be visible in the phenotype.
Elongation: The assembly of new DNA from template strands with the help of DNA polymerases.
Enzymes: Proteins responsible for catalyzing (accelerating) various biochemical reactions in cells.
Epigenetic profile: The methylation pattern throughout a genome—that is, which genes (and other genomic sites) are methylated and unmethylated.
Epigenetics: Changes in gene expression that do not result in a change of the underlying DNA sequence. These changes typically involve DNA methylation and histone modifications. These changes are reversible and can also be inherited by the next generation.
Euchromatin: Loosely coiled chromosomes found within the nucleus that are accessible for regulatory processing of DNA.
Eukaryote: Single-celled or multicelled organism characterized by a distinct nucleus, with each organelle surrounded by its own membrane.
Exon: Protein-coding segment of a gene.
Gametes: Haploid cells referred to as an egg and sperm that will fuse together during sexual reproduction to form a diploid organism.
Gene: Segment of DNA that contains protein-coding information and various regulatory (e.g., promoter) and noncoding (e.g., introns) regions.
Genetic recombination: A cellular process that occurs during meiosis I in which homologous chromosomes pair up and sister chromatids on different chromosomes physically swap genetic information.
Genome: All the genetic information of an organism.
Genotype: The combination of two alleles that code for or are associated with the same gene.
Genotyping: A molecular procedure that is performed to test for the presence of certain alleles or to discover new ones.
Germ cells: Specialized cells that form gametes (egg and sperm cells).
Haploid: Cell or organism with one set of chromosomes (n = 23).
Helicase: A protein that breaks the hydrogen bonds that hold double-stranded DNA together.
Heterozygous: Genotype that consists of two different alleles.
Histones: Proteins that DNA wraps around to assist with DNA organization within the nucleus.
Homologous chromosomes: A matching pair of chromosomes wherein one chromosome is maternally inherited and the other is paternally inherited.
Homozygous: Genotype that consists of two identical alleles.
Incomplete dominance: Heterozygous genotype that produces a phenotype that is a blend of both alleles.
Initiation: The recruitment of proteins to separate DNA strands and begin DNA replication.
Interphase: Preparatory period of the cell cycle when increased metabolic demand allows for DNA replication and doubling of the cell prior to cell division.
Introns: Segment of DNA that does not code for proteins.
Karyotyping: The microscopic procedure wherein the number of chromosomes in a cell is determined.
Lagging strand: DNA template strand that is opposite to the leading strand during DNA replication. This strand is created in several disconnected sections and other enzymes fill in the missing nucleotide gaps between these sections.
Leading strand: DNA template strand in which replication proceeds continuously.
Lipids: Fatty acid molecules that serve various purposes in the cell, including energy storage, cell signaling, and structure.
Meiosis: The process that gametes undergo to divide. The end of meiosis results in four haploid daughter cells.
Mendelian genetics: A classification given to phenotypic traits that are controlled by a single gene.
Messenger RNA (mRNA): RNA molecule that is transcribed from DNA. Its tri-nucleotide codons are “read” by a ribosome to build a protein.
Microarray technology: A genotyping procedure that utilizes a microarray chip, which is a collection of thousands of short nucleotide sequences attached to a solid surface that can probe genomic DNA.
Microbiome: The collective genomes of the community of microorganisms that humans have living inside of their bodies.
Mitochondrial DNA (mtDNA): Circular DNA segment found in mitochondria that is inherited maternally.
Mitochondrion: Specialized cellular organelle that is the site for energy production. It also has its own genome (mtDNA).
Mitosis: The process that somatic cells undergo to divide. The end of mitosis results in two diploid daughter cells.
Molecular anthropologists: Individuals who use molecular techniques (primarily genetics) to compare ancient and modern populations and to study living populations of humans and nonhuman primates.
Molecular geneticists: Biologists that study the structure and function of genes.
Mutation: A nucleotide sequence variation from the template DNA strand that can occur during replication. Mutations can also happen during recombination.
Next-generation sequencing: A genotyping technology that involves producing millions of nucleotide sequences (from a single DNA sample) that are then read with a sequencing machine. It can be used for analyzing entire genomes or specific regions and requires extensive program-based applications.
Nuclear envelope: A double-layered membrane that encircles the nucleus.
Nucleic acid: A complex structure (like DNA or RNA) that carries genetic information about a living organism.
Nucleotide: The basic structural component of nucleic acids, which includes DNA (A, T, C, and G) and RNA (A, U, C, and G).
Nucleus: Double-membrane cellular organelle that helps protect DNA and also regulates nuclear activities.
Organelle: A structure within a cell that performs specialized tasks that are essential for the cell. There are different types of organelles, each with its own function.
Pathogenic: A genetic mutation (i.e., allele) that has a harmful phenotypic disease-causing effect.
Pedigree: A diagram of family relationships that indicates which members may have or carry certain genetic and/or phenotypic traits.
Penetrance: The proportion of how often the possession of an allele results in an expected phenotype. Some alleles are more penetrant than others.
Phenotype: The physical appearance of a given trait.
Phospholipid bilayer: Two layers of lipids that form a barrier due to the properties of a hydrophilic (water-loving) head and a hydrophobic (water-repelling) tail.
Polygenic trait: A phenotype that is controlled by two or more genes.
Polymerase chain reaction (PCR): A molecular biology procedure that can make copies of genomic DNA segments. A small amount of DNA is used as a starting template and is then used to make millions of copies.
Prokaryote: A single-celled organism characterized by the lack of a nucleus and membrane-enclosed organelles.
Promoter: The region of a gene that initiates transcription. Transcription factors can bind and DNA methylation may occur at a promoter site, which can modify the transcriptional activities of a gene.
Protein: Chain of amino acids that folds into a three-dimensional structure that allows a cell to function in a variety of ways.
Protein synthesis: A multi-step process by which amino acids are strung together by RNA machinery read from a DNA template.
Recessive: Refers to an allele whose effect is not normally seen unless two copies are present in an individual’s genotype.
Ribonucleic acid (RNA): Single-stranded nucleic acid molecule.There are different RNAs found within cells and they perform a variety of functions, such as cell signaling and involvement in protein synthesis.
Ribosomal RNA (rRNA): A ribosome-bound molecule that is used to correctly assemble amino acids into proteins.
Ribosome: An organelle in the cell found in the cytoplasm or endoplasmic reticulum. It is responsible for reading mRNA and protein assemblage.
RNA polymerase: An enzyme that catalyzes the process of making RNA from a DNA template.
Sanger-sequencing: A process that involves the usage of fluorescently labeled nucleotides to visualize DNA (PCR fragments) at the nucleotide level.
Semi-conservative replication: DNA replication in which new DNA is replicated from an existing DNA template strand.
Sequencing: A molecular laboratory procedure that produces the order of nucleotide bases (i.e., sequences).
Sister chromatids: During DNA replication, sister chromatids are produced on the chromosome. In cell division, sister chromatids are pulled apart so that two cells can be formed. In meiosis, sister chromatids are also the sites of genetic recombination.
Somatic cells: Diploid cells that comprise body tissues and undergo mitosis for maintenance and repair of tissues.
Splicing: The process by which mature mRNAs are produced. Introns are removed (spliced) and exons are joined together.
Sugar phosphate backbone: A biochemical structural component of DNA. The “backbone” consists of deoxyribose sugars and phosphate molecules.
Telomere: A compound structure located at the ends of chromosomes to help protect the chromosomes from degradation after every round of cell division.
Termination: The halt of DNA replication activity that occurs when a DNA sequence “stop” codon is encountered.
Tissue: A cluster of cells that are morphologically similar and perform the same task.
Transcription: The process by which DNA nucleotides (within a gene) are copied, which results in a messenger RNA molecule.
Transcription factors: Proteins that bind to regulatory regions of genes (e.g., promoter) and increase or decrease the amount of transcriptional activity of a gene, including turning them “on” or “off.”
Transfer RNA (tRNA): RNA molecule involved in translation. Transfer RNA transports amino acids from the cell’s cytoplasm to a ribosome.
Translation: The process by which messenger RNA codons are read and amino acids are “chained together” to form proteins.
X-linked: Refers to a pattern of inheritance where the allele is located on the X or Y chromosome.
For Further Exploration
National Human Genome Research Institute
NOVA. 2018. Gene Sequencing Speeds Diagnosis of Deadly Newborn Diseases. NOVA, March 7, 2018. Accessed January 31, 2023. https://www.pbs.org/wgbh/nova/next/body/newborn-gene-sequencing/.
Zimmer, Carl. N.d. “Carl Zimmer’s Game of Genomes.” STATnews. Accessed January 31, 2023. https://www.statnews.com/feature/game-of-genomes/season-one/.
Illumina. 2016. “Illumina Sequencing by Synthesis.” YouTube.com, October 5, 2016. Accessed January 31, 2023. https://www.youtube.com/watch?v=fCd6B5HRaZ8.
References
Aartsma-Rus, Annemieke, Ieke B. Ginjaar, and Kate Bushby. 2016. “The Importance of Genetic Diagnosis for Duchenne Muscular Dystrophy.” Journal of Medical Genetics 53 (3): 145–151.
Acuna-Hidalgo, Rocio, Joris A. Veltman, and Alexander Hoischen. 2016. “New Insights into the Generation and Role of De Novo Mutations in Health and Disease.” Genome Biology 17 (241): 1–19.
Albert, Benjamin, Susanna Tomassetti, Yvonne Gloor, Daniel Dilg, Stefano Mattarocci, Slawomir Kubik, Lukas Hafner, and David Shore. 2019. "Sfp1 Regulates Transcriptional Networks Driving Cell Growth and Division through Multiple Promoter-Binding Modes." Genes & Development 33 (5–6): 288–293.
Almathen, Faisal, Haitham Elbir, Hussain Bahbahani, Joram Mwacharo, and Olivier Hanotte. 2018. “Polymorphisms in Mc1r and Asip Genes Are Associated with Coat Color Variation in the Arabian Camel.” Journal of Heredity 109 (6): 700–706.
Ballester, Leomar Y., Rajyalakshmi Luthra, Rashmi Kanagal-Shamanna, and Rajesh R. Singh. 2016. “Advances in Clinical Next-Generation Sequencing: Target Enrichment and Sequencing Technologies.” Expert Review of Molecular Diagnostics 16 (3): 357–372.
Baranovskiy, Andrey G., Vincent N. Duong, Nigar D. Babayeva, Yinbo Zhang, Youri I. Pavlov, Karen S. Anderson, and Tahir H. Tahirov. 2018. “Activity and Fidelity of Human DNA Polymerase Alpha Depend on Primer Structure.” Journal of Biological Chemistry 293 (18): 6824–6843.
Biswas, S., Ghosh, S., Das, S., & Maitra, S. (2021). Female Reproduction: At the Crossroads of Endocrine Disruptors and Epigenetics. Proceedings of the Zoological Society, 74(4), 532–545. https://doi.org/10.1007/s12595-021-00403-4
Brezina, Paulina R., Raymond Anchan, and William G. Kearns. 2016. “Preimplantation Genetic Testing for Aneuploidy: What Technology Should You Use and What Are the Differences?” Journal of Assisted Reproduction and Genetics 33 (7): 823–832.
Bultman, Scott J. 2017. “Interplay Between Diet, Gut Microbiota, Epigenetic Events, and Colorectal Cancer." Molecular Nutrition & Food Research 61 (1):1–12.
Cutting, Garry R. 2015. “Cystic Fibrosis Genetics: From Molecular Understanding to Clinical Application.” Nature Reviews Genetics 16 (1): 45–56.
D'Alessandro, Giuseppina., and Fabrizio d'Adda di Fagagna. 2017. “Transcription and DNA Damage: Holding Hands or Crossing Swords?” Journal of Molecular Biology 429 (21): 3215–3229.
De Craene, Johan-Owen, Dimitri L. Bertazzi, Séverine Bar, and Sylvie Friant. 2017. “Phosphoinositides, Major Actors in Membrane Trafficking and Lipid Signaling Pathways.” International Journal of Molecular Sciences 18 (3): 1–20.
Deng, Lian, and Shuhua Xu. 2018. “Adaptation of Human Skin Color in Various Populations.” Hereditas 155 (1): 1–12.
Dever, Thomas E., Terri G. Kinzy, and Graham D. Pavitt. 2016. “Mechanism and Regulation of Protein Synthesis in Saccharomyces Cerevisiae.” Genetics 203 (1): 65–107.
Eme, Laura, Anja Spang, Jonathan Lombard, Courtney W. Stairs, and Thijs J. G. Ettema. 2017. “Archaea and the Origin of Eukaryotes.” Nature Reviews Microbiology 15 (12): 711–723.
Gomez-Carballa, Alberto, Jacobo Pardo-Seco, Stefania Brandini, Alessandro Achilli, Ugo A. Perego, Michael D. Coble, Toni M. Diegoli, et al. 2018. “The Peopling of South America and the Trans-Andean Gene Flow of the First Settlers.” Genome Research 28 (6): 767–779.
Gvozdenov, Zlata, Janhavi Kolhe, and Brian C. Freeman. 2019. “The Nuclear and DNA-Associated Molecular Chaperone Network.” Cold Spring Harbor Perspectives in Biology 11 (10): a034009.
Harkins, Kelly M., and Anne C. Stone. 2015. “Ancient Pathogen Genomics: Insights into Timing and Adaptation.” Journal of Human Evolution 79: 137–149.
Jackson, Maria, Leah Marks, Gerhard H. W. May, and Joanna B. Wilson. 2018. “The Genetic Basis of Disease.” Essays in Biochemistry 62 (5): 643–723.
Lenormand, Thomas, Jan Engelstadter, Susan E. Johnston, Erik Wijnker, and Christopher R. Haag. 2016. “Evolutionary Mysteries in Meiosis.” Philosophical Transactions of the Royal Society B 371: 1–14.
Levy, Shawn E., and Richard M. Myers. 2016. “Advancements in Next-Generation Sequencing.” Annual Review of Genomics and Human Genetics 17: 95–115.
Lindo, John, Emilia Huerta-Sánchez, Shigeki Nakagome, Morten Rasmussen, Barbara Petzelt, Joycelynn Mitchell, Jerome S. Cybulski, et al. 2016. "A Time Transect of Exomes from a Native American Population Before and After European Contact." Nature Communications 7: 1–11. https://doi.org/10.1038/ncomms13175.
Lu, Mengfei, Cathryn M. Lewis, and Matthew Traylor. 2017. “Pharmacogenetic Testing through the Direct-to-Consumer Genetic Testing Company 23andme.” BMC Medical Genomics 10 (47): 1–8.
Ly, Lundi, Donovan Chan, Mahmoud Aarabi, Mylene Landry, Nathalie A. Behan, Amanda J. MacFarlane, and Jacquetta Trasler. 2017. “Intergenerational Impact of Paternal Lifetime Exposures to Both Folic Acid Deficiency and Supplementation on Reproductive Outcomes and Imprinted Gene Methylation.” Molecular Human Reproduction 23 (7): 461–477.
Ma, Wenxiu, Giancarlo Bonora, Joel B. Berletch, Xinxian Deng, William S. Noble, and Christine M. Disteche. 2018. “X-Chromosome Inactivation and Escape from X Inactivation in Mouse.” Methods in Molecular Biology 1861: 205–219.
Machiela, Mitchell J., Weiyin Zhou, Eric Karlins, Joshua N. Sampson, Neal D. Freedman, Qi Yang, Belynda Hicks, et al. 2016. “Female Chromosome X Mosaicism Is Age-Related and Preferentially Affects the Inactivated X Chromosome.” Nature Communications 7: 1–9. https://doi.org/10.1038/ncomms11843.
Mahdavi, Morteza, Mohammadreza Nassiri, Mohammad M. Kooshyar, Masoume Vakili-Azghandi, Amir Avan, Ryan Sandry, Suja Pillai, Alfred K. Lam, and Vinod Gopalan. 2019. “Hereditary Breast Cancer; Genetic Penetrance and Current Status with BRCA.” Journal of Cellular Physiology 234 (5): 5741–5750.
McDade, Thomas W., Calen P. Ryan, Meaghan J. Jones, Morgan K. Hoke, Judith Borja, Gregory E. Miller, Christopher W. Kuzawa, and Michael S. Kobor. 2019. “Genome-Wide Analysis of DNA Methylation in Relation to Socioeconomic Status During Development and Early Adulthood.” American Journal of Physical Anthropology 169 (1): 3–11.
Migeon, Barbara R. 2017. “Choosing the Active X: The Human Version of X Inactivation.” Trends in Genetics 33 (12): 899–909.
Myerowitz, Rachel. 1997. “Tay-Sachs Disease-Causing Mutations and Neutral Polymorphisms in the Hex A Gene.” Human Mutation 9 (3): 195–208.
National Institute of Environmental Health Sciences. (2024, July 22). Endocrine Disruptors. National Institute of Environmental Health Sciences; United States Government. https://www.niehs.nih.gov/health/topics/agents/endocrine
Onufriev, Alexey V., and Helmut Schiessel. 2019. “The Nucleosome: From Structure to Function through Physics.” Current Opinion in Structural Biology 56: 119–130.
Peedicayil J. (2006). Epigenetic therapy--a new development in pharmacology. The Indian journal of medical research, 123(1), 17–24.
Quillen, Ellen E., Heather L. Norton, Esteban J. Parra, Frida Lona-Durazo, Khai C. Ang, Florin M. Illiescu, Laurel N. Pearson, et al. 2019. “Shades of Complexity: New Perspectives on the Evolution and Genetic Architecture of Human Skin.” American Journal of Physical Anthropology 168 (67): 4–26.
Raspelli, Erica, and Roberta Fraschini. 2019. “Spindle Pole Power in Health and Disease.” Current Genetics 65 (4): 851–855.
Ravinet, M., R. Faria, R. K. Butlin, J. Galindo, N. Bierne, M. Rafajlovic, M. A. F. Noor, B. Mehlig, and A. M. Westram. 2017. “Interpreting the Genomic Landscape of Speciation: A Road Map for Finding Barriers to Gene Flow.” Journal of Evolutionary Biology 30 (8): 1450–1477.
Regev, Aviv, Sarah A. Teichmann, Eric S. Lander, Ido Amit, Christophe Benoist, Ewan Birney, Bernd Bodenmiller, et al. 2017. “The Human Cell Atlas.” Elife 6e27041: 1–30. https://doi.org/10.7554.eLife.27041.
Roberts, Andrea L., Nicole Gladish, Evan Gatev, Meaghan J. Jones, Ying Chen, Julia L. MacIsaac, Shelley S. Tworoger, et al. 2018. “Exposure to Childhood Abuse Is Associated with Human Sperm DNA Methylation.” Translational Psychiatry 8 (194): 1–11.
Roger, Andrew J., Sergio A. Muñoz-Gómez, and Ryoma Kamikawa. 2017. “The Origin and Diversification of Mitochondria.” Current Biology 27 (21): R1177–R1192. https://www.sciencedirect.com/science/article/pii/S096098221731179X?via%3Dihub#!
Ségurel, Laure, and Céline Bon. 2017. “On the Evolution of Lactase Persistence in Humans.” Annual Review of Genomics and Human Genetics 18: 297–319.
Sheth, Bhavisha P., and Vrinda S. Thaker. 2017. “DNA Barcoding and Traditional Taxonomy: An Integrated Approach for Biodiversity Conservation.” Genome 60 (7): 618–628.
Skloot, Rebecca. 2010. The Immortal Life of Henrietta Lacks. New York: Crown Publishing Group.
Snedeker, Jonathan, Matthew Wooten, and Xin Chen. 2017. “The Inherent Asymmetry of DNA Replication.” Annual Review of Cell and Developmental Biology 33: 291–318.
Sullivan-Pyke, Chantae, and Anuja Dokras. 2018. “Preimplantation Genetic Screening and Preimplantation Genetic Diagnosis.” Obstetrics and Gynecology Clinics of North America 45 (1): 113–125.
Sweeney, M. F., Hasan, N., Soto, A. M., & Sonnenschein, C. (2015). Environmental endocrine disruptors: Effects on the human male reproductive system. Reviews in Endocrine and Metabolic Disorders, 16(4), 341–357. https://doi.org/10.1007/s11154-016-9337-4
Szostak, Jack W. 2017. “The Narrow Road to the Deep Past: In Search of the Chemistry of the Origin of Life.” Angewandte Chemie International Edition 56 (37): 11037–11043.
Tessema, Mathewos, Ulrich Lehmann, and Hans Kreipe. 2004. “Cell Cycle and No End.” Virchows Archiv European Journal of Pathology 444 (4): 313–323.
Tishkoff, Sarah A., Floyd A. Reed, Alessia Ranciaro, Benjamin F. Voight, Courtney C. Babbitt, Jesse S. Silverman, Kweli Powell, et al. 2007. “Convergent Adaptation of Human Lactase Persistence in Africa and Europe.” Nature Genetics 39 (1): 31–40.
Visootsak, Jeannie, and John M. Graham, Jr. 2006. “Klinefelter Syndrome and Other Sex Chromosomal Aneuploidies.” Orphanet Journal of Rare Diseases 1:42. https://doi.org/10.1186/1750-1172-1-42.
Wolfe, George C., dir. 2017. The Immortal Life of Henrietta Lacks. HBO Films, April 22, 2017. TV Movie.
Yamamoto, Fumi-ichiro, Henrik Clausen, Thayer White, John Marken, and Sen-itiroh Hakomori. 1990. “Molecular Genetic Basis of the Histo-Blood Group ABO System.” Nature 345 (6272): 229–233.
Yu, X., Zhao, H., Wang, R., Chen, Y., Ouyang, X., Li, W., Sun, Y., & Peng, A. (2024). Cancer epigenetics: from laboratory studies and clinical trials to precision medicine. Cell Death Discovery, 10(1), 1–12. https://doi.org/10.1038/s41420-024-01803-z
Zlotogora, Joël. 2003. “Penetrance and Expressivity in the Molecular Age.” Genetics in Medicine 5 (5): 347–352.
Zorina-Lichtenwalter, Katerina, Ryan N. Lichtenwalter, Dima V. Zaykin, Marc Parisien, Simon Gravel, Andrey Bortsov, and Luda Diatchenko. 2019. “A Study in Scarlet: MC1R as the Main Predictor of Red Hair and Exemplar of the Flip-Flop Effect.” Human Molecular Genetics 28 (12): 2093-2106.
Zwart, Haeh. 2018. “In the Beginning Was the Genome: Genomics and the Bi-Textuality of Human Existence.” New Bioethics 24 (1): 26–43.

{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}